土壤水分胁迫对玉米形态发育及产量的影响
2
2004
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
4
2017
... 第二,根据气候资源特点,调整种植结构和种植制度以趋利避害.在半干旱地区通过实行“压夏扩秋”(即压缩春小麦、春玉米等春季作物种植比例)措施,扩大适应干旱能力较强且与降雨同季的马铃薯、大豆等秋季作物种植面积,应对干旱的季节变化特征.在以雨养农业为主的陇东黄土高原区,农业生产中稳定冬小麦种植面积,增加玉米面积,发展马铃薯、豆类、糜子、谷子等抗旱性较强的作物(邓振镛等,2007;张强等,2012b;肖国举和李裕,2012).合理安排和调整作物种植面积和布局,加强水热资源的合理开发利用和管理,变被动抗旱为主动抗旱,管好、用好当地水资源,充分利用大气降水(张强等,2012b;王润元等,2015;鲍文中和周广胜,2017). ...
... 第四,因地制宜推广保护性耕作措施,可同时起到保水、保土、培肥、增产的综合作用,如垄沟种植、垄沟径流集水、地膜覆盖、秸秆覆盖、免耕栽培等技术.采取多种形式的带状间作为中心的保护性耕作技术,缓解气候变暖加剧引起的水资源供求矛盾.推行麦类等条播作物与马铃薯、玉米、大豆等穴播作物或主粮作物与畜草合理间作、轮作等技术;合理套作,增加复种指数,提高耕地利用效率,如小麦套种马铃薯、小麦套种玉米等,可不同程度地提高作物产量,相当于在半干旱偏旱区的旱作农田上实现一年二熟(山仑,2011;张强等,2012a;张强等,2012b;肖国举和李裕,2012;鲍文中和周广胜,2017;Zhao et al.,2012;Zhao et al.,2014). ...
... 第五,改进施肥方式和用量,测土施肥,配合深施、混施等施肥方式,提高肥效和作物对营养元素的利用率,如玉米施氮增效不仅可以获得较高产量、水分和氮素利用效率,还能降低硝态氮的残留量,有效减缓土壤污染(肖国举和李裕,2012;Qiang et al.,2019a).冬小麦测土施肥,可用于诊断其氮营养状况,提高水分利用效率、增加叶面积指数和干物质积累(肖国举和李裕,2012;Qiang et al.,2019b).未来半干旱地区的小麦生产,可视温度、CO2浓度升高情况适量采用低磷肥、低氮肥投入,这将更经济、适宜(王润元等,2015;肖国举和李裕,2012;鲍文中和周广胜,2017). ...
... 第六,加强农业干旱监测预测预警,强化防旱减灾应对准备.在出现旱情的重点地区加强土壤墒情监测和定点调查,构建区域信息共享平台,及时掌握旱情发展蔓延动态;加快构建农业农村气象灾害预警信息发布系统,搭建多种渠道的气象服务信息传播平台,建立气象灾害风险评估体系,研发基于影响的农业干旱监测预测预警技术,针对农业生产中重要环节和农作物生长发育的水分需求,提升农业防旱减灾服务能力,定期发布土壤墒情监测公报、干旱监测公报、农业气象旬(周)报等农业气象服务产品(姚国章和袁敏,2010;张强等,2014;周广胜等,2016).总之,在确保国家和区域(或省、市、区)粮、棉、油需求安全前提下,充分利用水热资源优势,压缩高耗水作物和品种的种植面积,实行农业补贴政策,实现农业经济和水资源安全协调发展(鲍文中和周广胜,2017). ...
干旱胁迫对夏玉米叶片光合及叶绿素荧光的影响
2
2008
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
气候变化背景下中国重大农业气象灾害预测预警技术研究
1
2012
... 2006年,美国开始建设协调综合的国家干旱早期预警系统(Drought Early Warning System, DEWS),随后又建成了国家综合干旱信息系统(National Integrated Drought Information System,NIDIS);2007年,地球观测部计划未来建立全球干旱早期预警系统(Global Drought Early Warning System,GDEWS).美国利用MODIS(Moderate Resolution Imaging Spectroradiometer)产品监测全球干旱及旱灾造成的作物减产分布,用户可通过网络界面查询预先定置区域的数据并分析农作物长势和旱情(范锦龙等,2014).此外,美国国家干旱减灾中心(National Drought Mitigation Center,NDMC)联合美国农业部(The United States Department of Agriculture,USDA)等相关部门研发包括气象指标、土壤墒情指标的干旱监测系统(张晓煜等,2011;纪瑞鹏等,2019).中国风云卫星也具有监测全球干旱的能力,农业监测是其一个关键的应用领域,如FY-3A和FY-3B携带的可见光近红外扫描辐射计(Visible Infrared Radiometer,VIRR)和中分辨率成像仪(Medium Resolution Spectral Imager,MERSI)是农业监测的关键传感器,可监测不同尺度干旱灾害的发生发展(张晓煜等,2011;范锦龙等,2014).此外,中国国家气候中心也开展了干旱监测、预测预警和影响评估业务,发布全球旱涝指数、中国气象干旱综合指数、土壤相对湿度等系列干旱产品和公告,为抗旱减灾提供了重要的决策服务信息(范锦龙等,2014).在国家科技部支持下,气象、农业等部门联合运用气象灾害预测预报技术进行重大农业气象灾害监测预测和预警技术的攻关研发(陈德亮,2012;纪瑞鹏等,2019). ...
新时代防汛防旱信息化能力提升思考
1
2020
... 农业防旱减灾是一项复杂的社会系统工程,涉及旱灾发生前的监测、预报、预警和防御,以及灾害发生时抗灾、灾后救灾和重建等多个方面.依照“工程措施与非工程措施相结合”的原则,在加强水利工程设施建设的同时,还需强化政策法规、监测预警、旱中保障等抗旱非工程措施,构建旱情监测预警和决策系统,提高抗旱减灾工作的科学化、现代化和规范化水平(姚国章和袁敏,2010;周广胜等,2016).鉴于旱灾对农业生产影响较重,急需建立一个科学有效的农业旱灾保障机制,但目前我国这种保障机制仍较滞后,相应的农业保险等项目发展缓慢(姚国章和袁敏,2010;周广胜等,2016).我国在干旱灾害管理方面的法律法规和政策尚有待健全,需借鉴欧美等发达国家经验,由被动的应急抗旱向主动的防旱模式转变(毛晓华,2020;屈艳萍等,2013).同时,我国在各类干旱(包括气象、农业、水文等)监测方面已开展了大量研究工作,但缺乏各部门间的有效协作机制,即气象、水文、农情、工业取水用水和相应供水部门之间的信息共享和联防联动.干旱整体评价指标构建上还需借鉴国外的干旱综合评价指标体系(康天军和李军波,2018;陈滢,2020).未来,我国还需制定更为详细的干旱灾害防御规划,让公众参与到干旱防灾、减灾工作中,构建全民参与的全社会防灾减灾新格局. ...
干旱气候变化对甘肃省干旱灾害的影响及防旱减灾技术的研究
3
2007
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 图6是1980年以来全国9大地区重旱以上灾害发生频率.可以看出,1980年以后我国北方的内蒙古、西北地区、东北地区和黄淮海地区干旱灾害发生频繁且较为严重,重旱以上发生频率分别为45.0%、40.0%、35.0%和25.0%,受灾率和成灾率均超过15.0%和10.0%(倪深海等,2022).从干旱发生范围来看,1980年以前发生重旱以上的区域波及10个省(市、区),而1980年以后扩增到16个省(市、区)(倪深海等,2022).受气候变暖影响,近50 a来西北地区干旱频率、强度和受灾面积增加,损失加重.其中,春旱、秋旱频次增加,夏旱频次减少,春旱、秋旱多于夏旱,特、重旱多出现在春季,主要发生在西北地区东部,尤其是甘肃河东地区,如1995年干旱最严重,受灾面积208.7万hm2,成灾面积170.8万hm2,成灾率为45%,粮食减产150万t;其次是2000年,成灾率达34.9%,粮食减产135万t(邓振镛等,2007).预计未来一段时间内,如果不采取有效应对措施,在气候变暖背景下到2030年中国种植业生产力总体可能下降5%~10%,其中小麦、水稻和玉米三大作物均以降低为主,2050年后受影响会更大(IPCC,2022). ...
... 第二,根据气候资源特点,调整种植结构和种植制度以趋利避害.在半干旱地区通过实行“压夏扩秋”(即压缩春小麦、春玉米等春季作物种植比例)措施,扩大适应干旱能力较强且与降雨同季的马铃薯、大豆等秋季作物种植面积,应对干旱的季节变化特征.在以雨养农业为主的陇东黄土高原区,农业生产中稳定冬小麦种植面积,增加玉米面积,发展马铃薯、豆类、糜子、谷子等抗旱性较强的作物(邓振镛等,2007;张强等,2012b;肖国举和李裕,2012).合理安排和调整作物种植面积和布局,加强水热资源的合理开发利用和管理,变被动抗旱为主动抗旱,管好、用好当地水资源,充分利用大气降水(张强等,2012b;王润元等,2015;鲍文中和周广胜,2017). ...
全球干旱卫星监测计划
5
2014
... 针对农业干旱监测预警和风险评估,国内外组织和学者相继开展了一系列研发工作,如2007年以来,作为政府间的国际组织,地球观测组织(Group on Earth Observations,GEO)推动了全球农业监测计划(Global Agricultural Monitoring,GEO-GLAM),就农业干旱监测进行了大量探索工作,该计划支持发展和改进农业监测系统,旨在利用地球观测数据开展农业生产监测预警、农业土地利用变化观测、农业气象条件预测等活动,来加强全球粮食产量监测评估及对存在粮食安全风险国家的监测,增强全球农业监测能力和监测系统的建设.源于该计划的支持,我国自主研发的全球农情遥感监测系统(CropWatch云平台)已成为全球主要的3个农情监测系统之一,中国作为首个推出粮食遥感监测云服务的国家,已为全球超过170个国家和地区提供独立的农情信息服务,提高了粮食生产信息的透明度(范锦龙等,2014;纪瑞鹏等,2019; https://edu.yunnan.cn/system/2022/09/09/032270206.shtml).此外,GEO也推动了全球综合地球观测系统(Global Earth Observation System of Systems, GEO-SS),主要协调地球观测数据的广泛应用及提高应用能力来促进可持续农业、风险评估、粮食安全、市场效率等,通过发布天气预报和极端天气事件早期预警、中长期气候预测等信息支持农业可持续管理(范锦龙等,2014). ...
... ).此外,GEO也推动了全球综合地球观测系统(Global Earth Observation System of Systems, GEO-SS),主要协调地球观测数据的广泛应用及提高应用能力来促进可持续农业、风险评估、粮食安全、市场效率等,通过发布天气预报和极端天气事件早期预警、中长期气候预测等信息支持农业可持续管理(范锦龙等,2014). ...
... 2006年,美国开始建设协调综合的国家干旱早期预警系统(Drought Early Warning System, DEWS),随后又建成了国家综合干旱信息系统(National Integrated Drought Information System,NIDIS);2007年,地球观测部计划未来建立全球干旱早期预警系统(Global Drought Early Warning System,GDEWS).美国利用MODIS(Moderate Resolution Imaging Spectroradiometer)产品监测全球干旱及旱灾造成的作物减产分布,用户可通过网络界面查询预先定置区域的数据并分析农作物长势和旱情(范锦龙等,2014).此外,美国国家干旱减灾中心(National Drought Mitigation Center,NDMC)联合美国农业部(The United States Department of Agriculture,USDA)等相关部门研发包括气象指标、土壤墒情指标的干旱监测系统(张晓煜等,2011;纪瑞鹏等,2019).中国风云卫星也具有监测全球干旱的能力,农业监测是其一个关键的应用领域,如FY-3A和FY-3B携带的可见光近红外扫描辐射计(Visible Infrared Radiometer,VIRR)和中分辨率成像仪(Medium Resolution Spectral Imager,MERSI)是农业监测的关键传感器,可监测不同尺度干旱灾害的发生发展(张晓煜等,2011;范锦龙等,2014).此外,中国国家气候中心也开展了干旱监测、预测预警和影响评估业务,发布全球旱涝指数、中国气象干旱综合指数、土壤相对湿度等系列干旱产品和公告,为抗旱减灾提供了重要的决策服务信息(范锦龙等,2014).在国家科技部支持下,气象、农业等部门联合运用气象灾害预测预报技术进行重大农业气象灾害监测预测和预警技术的攻关研发(陈德亮,2012;纪瑞鹏等,2019). ...
... ;范锦龙等,2014).此外,中国国家气候中心也开展了干旱监测、预测预警和影响评估业务,发布全球旱涝指数、中国气象干旱综合指数、土壤相对湿度等系列干旱产品和公告,为抗旱减灾提供了重要的决策服务信息(范锦龙等,2014).在国家科技部支持下,气象、农业等部门联合运用气象灾害预测预报技术进行重大农业气象灾害监测预测和预警技术的攻关研发(陈德亮,2012;纪瑞鹏等,2019). ...
... ).此外,中国国家气候中心也开展了干旱监测、预测预警和影响评估业务,发布全球旱涝指数、中国气象干旱综合指数、土壤相对湿度等系列干旱产品和公告,为抗旱减灾提供了重要的决策服务信息(范锦龙等,2014).在国家科技部支持下,气象、农业等部门联合运用气象灾害预测预报技术进行重大农业气象灾害监测预测和预警技术的攻关研发(陈德亮,2012;纪瑞鹏等,2019). ...
农业干旱的成因、指标、时空分布和防旱抗旱对策
1
1995
... 农业干旱应对主要包括农业干旱的减轻和防御两方面,农业干旱的发生、发展与气象干旱、水文干旱等其他类型干旱紧密相关(吴杰峰等,2017),因此在应对和防御农业干旱时,不仅要关注农业干旱,还要加强其他类型干旱的监测和管理.农业干旱的减轻措施是指用来抵御干旱灾害的不利影响、环境恶化以及技术风险的工程措施(如基础设施、修筑堤坝、工程建设等)或非工程措施(如政策法规、政府决策、公众意识等).农业干旱的防御措施是指在干旱灾害发生前制定的政策规划、制度以及灾害的监测、预测或预警,以确保灾害发生时各方面的协调与有效响应(冯定原和邱新法,1995;陆亚龙和肖功建,2001).粮食生产对干旱的减轻、防御和适应技术对策见图8. ...
4
2015
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 在农业干旱监测中,农业干旱指标主要有气象指标、土壤墒情指标、作物生理生态指标和一些其他指标.气象指标包括降水距平百分数、连续无有效降水日数、湿度指标和Z指数等(孙荣强,1994;姚玉璧等,2013;国家气象中心等,2015;Zhang et al.,2009).土壤墒情指标包括土壤相对湿度、土壤有效水分存储量、土壤水分旱灾指数等(国家气象中心等,2015;Hollinger et al.,1993;Dai et al.,2004).作物生理生态指标主要是一些基于植株生长和光合特性的生理参数,用来衡量植株水分亏缺状况,如光合速率、蒸腾速率、气孔导度、复水后的光合恢复、叶水势、茎秆水势、叶绿素、叶片相对含水量、叶片扩展速率、作物水分胁迫指数、冠层温度、块茎碳同位素鉴别等(姚玉璧等,2013;张强等,2014;赵鸿等,2018;纪瑞鹏等,2019;Jefferies,1993;Medrano et al.,2002;Zakaluk and Ranjan,2006;Ricardo,2012;Ramirez et al.,2016),这些基于植株的监测指标更加方便、适时,但有些地区由于没有专用测量仪器而导致部分指标不能广泛应用. ...
... ).土壤墒情指标包括土壤相对湿度、土壤有效水分存储量、土壤水分旱灾指数等(国家气象中心等,2015;Hollinger et al.,1993;Dai et al.,2004).作物生理生态指标主要是一些基于植株生长和光合特性的生理参数,用来衡量植株水分亏缺状况,如光合速率、蒸腾速率、气孔导度、复水后的光合恢复、叶水势、茎秆水势、叶绿素、叶片相对含水量、叶片扩展速率、作物水分胁迫指数、冠层温度、块茎碳同位素鉴别等(姚玉璧等,2013;张强等,2014;赵鸿等,2018;纪瑞鹏等,2019;Jefferies,1993;Medrano et al.,2002;Zakaluk and Ranjan,2006;Ricardo,2012;Ramirez et al.,2016),这些基于植株的监测指标更加方便、适时,但有些地区由于没有专用测量仪器而导致部分指标不能广泛应用. ...
... 此外,还有一些重要的监测指标如帕尔默干旱指数、作物水分亏缺指数、作物水分指数、农田与作物形态农业干旱指标、作物特征旱灾指数、减产率农业干旱指数、综合农业干旱指数、区域性农业干旱指标以及农业干旱过程的确定和评估等(Palmer,1968;李柏贞和周广胜,2014;国家气象中心等,2015).其中,帕尔默干旱指数是目前国际上应用最广泛的干旱指标,它引入了水量平衡概念,包含降水量、蒸散量、径流量和土壤水分储存量等,用于监测以月为时间尺度的干旱(表1).该指数不仅考虑了当前的水分条件,还考虑了前期水分状况及干旱持续时间等,具有较好的时空可比性,能够较好地监测评估较长时期的干旱,同时也是衡量土壤湿度和确定干旱开始和终止时间最有效的评估方法,但该指标考虑的因素较多,对资料要求较高,无法实现作物对水分亏缺响应的逐日监测. ...
以色列的抗旱措施
2
2002
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 以色列国家面对水资源严重贫乏的现状,首先制定了《水法》,颁布全国水资源管理和用水政策,实施工农业用水配额,大力推行农业节水灌溉、废水处理灌溉、微咸水灌溉等措施,将每一滴水都用到了极致(韩萍,2002).瑞典、荷兰、德国等欧洲国家利用人工补给含水层来增加水资源量,极力发挥地下水库调节作用以抵御干旱风险(https://www.docin.com/p-2012740929.html).欧盟也制定了一系列旱灾风险管理战略,从水资源危机管理向干旱风险管理转变(https://world.qianlong.com/2022/0830/7575142.shtml).埃塞俄比亚、肯尼亚、索马里等非洲国家则通过积极引进抗旱作物和抗旱技术应对农业干旱造成的粮食短缺问题(吴爱民,2011),如为缓解干旱造成的粮食短缺,引进了具有耐旱性的玉米新品种,水稻高杆品种变为矮杆,并辅配农药和农业机械,解决了19个发展中国家的粮食自给问题(https://world.qianlong.com/2022/0830/7575142.shtml). ...
作物对干旱胁迫的响应过程与早期识别技术研究进展
6
2019
... 2022年气候异常,北半球遭遇了大范围、史无前例的严重干旱,北美洲、欧洲、地中海地区、东北非地区以及中国南方地区都出现破纪录的极端酷热天气,许多地区经历了极端干旱(图2),对农业生产产生较大影响,大部分农作物有一定程度的减产.2022年9月联合国粮农组织发布的月报,将2022年全球谷物产量预测从7月初预测的27.92亿t下调至27.74亿t,比2021年预估产量低1.4%(https://xueqiu.com/4587623715/231014661).1964—2007年极端天气灾害造成全球粮食作物大面积减产,尤其是玉米、水稻、小麦,其中干旱和极端高温导致各国粮食减产9%~10%,发达国家的粮食产量损失比发展中国家高出8%~11%(纪瑞鹏等,2019;Lesk et al.,2016;Mirzabaev et al.,2023) ...
... 在农业干旱监测中,农业干旱指标主要有气象指标、土壤墒情指标、作物生理生态指标和一些其他指标.气象指标包括降水距平百分数、连续无有效降水日数、湿度指标和Z指数等(孙荣强,1994;姚玉璧等,2013;国家气象中心等,2015;Zhang et al.,2009).土壤墒情指标包括土壤相对湿度、土壤有效水分存储量、土壤水分旱灾指数等(国家气象中心等,2015;Hollinger et al.,1993;Dai et al.,2004).作物生理生态指标主要是一些基于植株生长和光合特性的生理参数,用来衡量植株水分亏缺状况,如光合速率、蒸腾速率、气孔导度、复水后的光合恢复、叶水势、茎秆水势、叶绿素、叶片相对含水量、叶片扩展速率、作物水分胁迫指数、冠层温度、块茎碳同位素鉴别等(姚玉璧等,2013;张强等,2014;赵鸿等,2018;纪瑞鹏等,2019;Jefferies,1993;Medrano et al.,2002;Zakaluk and Ranjan,2006;Ricardo,2012;Ramirez et al.,2016),这些基于植株的监测指标更加方便、适时,但有些地区由于没有专用测量仪器而导致部分指标不能广泛应用. ...
... 针对农业干旱监测预警和风险评估,国内外组织和学者相继开展了一系列研发工作,如2007年以来,作为政府间的国际组织,地球观测组织(Group on Earth Observations,GEO)推动了全球农业监测计划(Global Agricultural Monitoring,GEO-GLAM),就农业干旱监测进行了大量探索工作,该计划支持发展和改进农业监测系统,旨在利用地球观测数据开展农业生产监测预警、农业土地利用变化观测、农业气象条件预测等活动,来加强全球粮食产量监测评估及对存在粮食安全风险国家的监测,增强全球农业监测能力和监测系统的建设.源于该计划的支持,我国自主研发的全球农情遥感监测系统(CropWatch云平台)已成为全球主要的3个农情监测系统之一,中国作为首个推出粮食遥感监测云服务的国家,已为全球超过170个国家和地区提供独立的农情信息服务,提高了粮食生产信息的透明度(范锦龙等,2014;纪瑞鹏等,2019; https://edu.yunnan.cn/system/2022/09/09/032270206.shtml).此外,GEO也推动了全球综合地球观测系统(Global Earth Observation System of Systems, GEO-SS),主要协调地球观测数据的广泛应用及提高应用能力来促进可持续农业、风险评估、粮食安全、市场效率等,通过发布天气预报和极端天气事件早期预警、中长期气候预测等信息支持农业可持续管理(范锦龙等,2014). ...
... 2006年,美国开始建设协调综合的国家干旱早期预警系统(Drought Early Warning System, DEWS),随后又建成了国家综合干旱信息系统(National Integrated Drought Information System,NIDIS);2007年,地球观测部计划未来建立全球干旱早期预警系统(Global Drought Early Warning System,GDEWS).美国利用MODIS(Moderate Resolution Imaging Spectroradiometer)产品监测全球干旱及旱灾造成的作物减产分布,用户可通过网络界面查询预先定置区域的数据并分析农作物长势和旱情(范锦龙等,2014).此外,美国国家干旱减灾中心(National Drought Mitigation Center,NDMC)联合美国农业部(The United States Department of Agriculture,USDA)等相关部门研发包括气象指标、土壤墒情指标的干旱监测系统(张晓煜等,2011;纪瑞鹏等,2019).中国风云卫星也具有监测全球干旱的能力,农业监测是其一个关键的应用领域,如FY-3A和FY-3B携带的可见光近红外扫描辐射计(Visible Infrared Radiometer,VIRR)和中分辨率成像仪(Medium Resolution Spectral Imager,MERSI)是农业监测的关键传感器,可监测不同尺度干旱灾害的发生发展(张晓煜等,2011;范锦龙等,2014).此外,中国国家气候中心也开展了干旱监测、预测预警和影响评估业务,发布全球旱涝指数、中国气象干旱综合指数、土壤相对湿度等系列干旱产品和公告,为抗旱减灾提供了重要的决策服务信息(范锦龙等,2014).在国家科技部支持下,气象、农业等部门联合运用气象灾害预测预报技术进行重大农业气象灾害监测预测和预警技术的攻关研发(陈德亮,2012;纪瑞鹏等,2019). ...
... ;纪瑞鹏等,2019). ...
... 卫星遥感、地面遥感等探测技术的发展,为农作物旱情监测开辟了一条新途径.目前,国内外学者相继研发了多个农业干旱遥感监测指数,主要有以下四大类:(1)土壤水分指数,如表观热惯量(Apparent Thermal Inertia,ATI)、微波反演土壤水分等;(2)作物形态及绿度指数,如归一化植被指数(Normalized Difference Vegetation Index,NDVI)、植被状态指数(Vegetation Condition Index,VCI)、标准植被指数(Standardized Vegetation Index,SVI)等;(3)冠层温度指数,包括归一化温度指数(Normalized Difference Temperature Index,NDTI)、作物水分胁迫指数(Crop Water Stress Index,CWSI)、温度植被干旱指数(Temperature Vegetation Dryness Index,TVDI)、植被供水指数(Vegetation Supply Water Index,VSWI)等;(4)植被水分指数,包括全球植被水分指数(Global Vegetation Moisture Index,GVMI)、归一化差异水分指数(Normalized Difference Water Index,NDWI)等.这些指数各有优缺点,基于作物光谱信息,结合地面站点观测数据,可对大范围农作物干旱进行监测和评估,取得了较好的效果(李柏贞和周广胜,2014;纪瑞鹏等,2019;Anderson,2011). ...
我国五大粮食主产区农业干旱态势综合研究
2
2014
... 我国是一个农业大国,也是旱灾频发的国家,旱灾影响范围广、历时长、危害重.据统计,我国每年因干旱受灾的农田面积多达2 600万hm2,粮食减产高达50%~75%(姚玉璧等,2013;张强等,2014).近年来,受气候变化和水资源短缺的影响,我国农作物受旱、成灾和绝收面积不断扩大,在北方干旱形势依然严峻的情况下南方干旱出现明显增加和加重趋势,新旱区逐渐扩展,局部性、区域性干旱灾害频频出现,损失巨大(张强等,2014;康蕾和张红旗,2014;倪深海等,2022;张强,2022),严重威胁着国家的粮食安全和生态安全(王春乙,2007;张强等,2012a;张强等,2012b;肖国举和李裕,2012;康蕾和张红旗,2014;周广胜等,2016;Fu et al.,2013),已成为制约社会经济可持续发展的最主要因素,给我国农业生产和抗旱减灾工作带来前所未有的挑战和困难(吕娟等,2011;张强等,2014;Huang et al.,2016). ...
... ;康蕾和张红旗,2014;周广胜等,2016;Fu et al.,2013),已成为制约社会经济可持续发展的最主要因素,给我国农业生产和抗旱减灾工作带来前所未有的挑战和困难(吕娟等,2011;张强等,2014;Huang et al.,2016). ...
小麦内源激素脱落酸研究进展
1
1997
... 植物生长调节剂(外用)和植物激素(内用)是影响植物生理过程的物质,其浓度很低时起调节作用,如脱落酸、乙烯、生长素、赤霉素、细胞分裂素等(Morgan,1991;康书江等,1997).在分子和生化水平上,植物通过在转录和蛋白表达过程中改变调节反应来感知和响应干旱胁迫,从而影响生化途径和代谢过程,进而影响生理和发育过程(Mane et al.,2008;Vasquez-Robinet et al.,2008;Zinselmeier et al.,1995).在土壤干旱条件下植物通常会产生脱落酸(Abscisic Acid,ABA),它是一种主要的化学根-芽胁迫信号(Davies and Zhang,1991),是通过激活对干旱响应的基因表达而表征细胞反应的一个关键信号(Chaves et al.,2003).ABA含量可作为植物抗旱性的评价指标之一,其与籽粒的生长速率有关.当土壤干旱时失水的根系产生根源信号ABA,通过木质部输送到地上部调节气孔开闭(Davies and Zhang,1991),作物根冠、叶片、花、籽粒生长进程中ABA含量明显上升(赵鸿等,2008;张玉书等,2012;Obidiegwu et al,2015),且随着干旱胁迫程度的加剧而增加,如受到水分胁迫时小麦籽粒生长减慢、结实率降低,玉米胚乳细胞分化率降低.乙烯(Ethylene)也是干旱胁迫下植物体产生的激素,在干旱感知时乙烯通过蒸腾流将其他激素信号与ABA一起发送到嫩芽中.干旱引发的乙烯释放过量,会引起小麦、玉米等籽粒产量下降(赵鸿等,2008;Obidiegwu et al.,2015). ...
我国防旱抗旱法制体系的完善——基于政策指引及比较法的进路
1
2018
... 农业防旱减灾是一项复杂的社会系统工程,涉及旱灾发生前的监测、预报、预警和防御,以及灾害发生时抗灾、灾后救灾和重建等多个方面.依照“工程措施与非工程措施相结合”的原则,在加强水利工程设施建设的同时,还需强化政策法规、监测预警、旱中保障等抗旱非工程措施,构建旱情监测预警和决策系统,提高抗旱减灾工作的科学化、现代化和规范化水平(姚国章和袁敏,2010;周广胜等,2016).鉴于旱灾对农业生产影响较重,急需建立一个科学有效的农业旱灾保障机制,但目前我国这种保障机制仍较滞后,相应的农业保险等项目发展缓慢(姚国章和袁敏,2010;周广胜等,2016).我国在干旱灾害管理方面的法律法规和政策尚有待健全,需借鉴欧美等发达国家经验,由被动的应急抗旱向主动的防旱模式转变(毛晓华,2020;屈艳萍等,2013).同时,我国在各类干旱(包括气象、农业、水文等)监测方面已开展了大量研究工作,但缺乏各部门间的有效协作机制,即气象、水文、农情、工业取水用水和相应供水部门之间的信息共享和联防联动.干旱整体评价指标构建上还需借鉴国外的干旱综合评价指标体系(康天军和李军波,2018;陈滢,2020).未来,我国还需制定更为详细的干旱灾害防御规划,让公众参与到干旱防灾、减灾工作中,构建全民参与的全社会防灾减灾新格局. ...
半干旱区黑膜覆盖对马铃薯光合特性及产量的影响
1
2017
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
干旱指标研究进展
3
2014
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 此外,还有一些重要的监测指标如帕尔默干旱指数、作物水分亏缺指数、作物水分指数、农田与作物形态农业干旱指标、作物特征旱灾指数、减产率农业干旱指数、综合农业干旱指数、区域性农业干旱指标以及农业干旱过程的确定和评估等(Palmer,1968;李柏贞和周广胜,2014;国家气象中心等,2015).其中,帕尔默干旱指数是目前国际上应用最广泛的干旱指标,它引入了水量平衡概念,包含降水量、蒸散量、径流量和土壤水分储存量等,用于监测以月为时间尺度的干旱(表1).该指数不仅考虑了当前的水分条件,还考虑了前期水分状况及干旱持续时间等,具有较好的时空可比性,能够较好地监测评估较长时期的干旱,同时也是衡量土壤湿度和确定干旱开始和终止时间最有效的评估方法,但该指标考虑的因素较多,对资料要求较高,无法实现作物对水分亏缺响应的逐日监测. ...
... 卫星遥感、地面遥感等探测技术的发展,为农作物旱情监测开辟了一条新途径.目前,国内外学者相继研发了多个农业干旱遥感监测指数,主要有以下四大类:(1)土壤水分指数,如表观热惯量(Apparent Thermal Inertia,ATI)、微波反演土壤水分等;(2)作物形态及绿度指数,如归一化植被指数(Normalized Difference Vegetation Index,NDVI)、植被状态指数(Vegetation Condition Index,VCI)、标准植被指数(Standardized Vegetation Index,SVI)等;(3)冠层温度指数,包括归一化温度指数(Normalized Difference Temperature Index,NDTI)、作物水分胁迫指数(Crop Water Stress Index,CWSI)、温度植被干旱指数(Temperature Vegetation Dryness Index,TVDI)、植被供水指数(Vegetation Supply Water Index,VSWI)等;(4)植被水分指数,包括全球植被水分指数(Global Vegetation Moisture Index,GVMI)、归一化差异水分指数(Normalized Difference Water Index,NDWI)等.这些指数各有优缺点,基于作物光谱信息,结合地面站点观测数据,可对大范围农作物干旱进行监测和评估,取得了较好的效果(李柏贞和周广胜,2014;纪瑞鹏等,2019;Anderson,2011). ...
中亚地区干旱变化及其影响分析
1
2022
... 受全球气候变化和经济快速发展的影响,全球干旱呈现多发、加重趋势,中亚地区65%的区域表现为干旱程度加剧、强度持续增强,2022年全球极端干旱问题尤为突出(李稚等,2022;张强,2022;Metz et al.,2007;Li et al.,2015;IPCC,2022).随着全球气候变暖,水循环加快,陆地蒸散发和储水量减少,干旱半干旱区不断扩张,干旱持续加剧,干旱风险进一步加大,且存在较大的区域差异(张强等,2017;Salinger et al.,2005;Cook et al.,2007;Metz et al.,2007;Huang et al.,2016).联合国《2022年全球干旱数字》报告指出,自2000年以来全球干旱次数增加了29%,全球处于高温和干旱管理的“十字路口”(UNCCD, 2022).预计到2100年,极端干旱地区的干旱次数和强度将增加1%~30%(IPCC,2007). ...
基于农业干旱参考指数的西南地区玉米干旱时空变化分析
2
2014
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
2
2001
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 农业干旱应对主要包括农业干旱的减轻和防御两方面,农业干旱的发生、发展与气象干旱、水文干旱等其他类型干旱紧密相关(吴杰峰等,2017),因此在应对和防御农业干旱时,不仅要关注农业干旱,还要加强其他类型干旱的监测和管理.农业干旱的减轻措施是指用来抵御干旱灾害的不利影响、环境恶化以及技术风险的工程措施(如基础设施、修筑堤坝、工程建设等)或非工程措施(如政策法规、政府决策、公众意识等).农业干旱的防御措施是指在干旱灾害发生前制定的政策规划、制度以及灾害的监测、预测或预警,以确保灾害发生时各方面的协调与有效响应(冯定原和邱新法,1995;陆亚龙和肖功建,2001).粮食生产对干旱的减轻、防御和适应技术对策见图8. ...
21世纪以来我国干旱灾害特点及成因分析
2
2011
... 干旱灾害(简称“旱灾”),是指一段时间内降水量比常年明显偏少,造成某一地区经济社会活动(尤其是农业生产)和人类生活遭受较大危害的现象,普遍存在于世界各地,频繁发生于不同时期(Wilhite,2000;Li et al.,2015).旱灾是气象干旱、农业干旱、水文干旱、社会经济干旱和生态干旱之间的一种链状传递和转化过程,一个完整的干旱事件包括干旱历时、严重程度和平均强度(姚玉璧等,2013;张强等,2014;Heffernan,2013).全球每年因干旱导致的经济损失达60~80亿美元,2021年甚至高达121亿美元(American Meteorological Society, 1997;应急管理部-教育部减灾与应急管理研究院等,2022),远高于其他气象灾害造成的损失(张强等,2014;Wilhite,2000),受到高度关注(王春乙,2007;吕娟等,2011;Turner et al., 2011;Gupta et al.,2020). ...
... 我国是一个农业大国,也是旱灾频发的国家,旱灾影响范围广、历时长、危害重.据统计,我国每年因干旱受灾的农田面积多达2 600万hm2,粮食减产高达50%~75%(姚玉璧等,2013;张强等,2014).近年来,受气候变化和水资源短缺的影响,我国农作物受旱、成灾和绝收面积不断扩大,在北方干旱形势依然严峻的情况下南方干旱出现明显增加和加重趋势,新旱区逐渐扩展,局部性、区域性干旱灾害频频出现,损失巨大(张强等,2014;康蕾和张红旗,2014;倪深海等,2022;张强,2022),严重威胁着国家的粮食安全和生态安全(王春乙,2007;张强等,2012a;张强等,2012b;肖国举和李裕,2012;康蕾和张红旗,2014;周广胜等,2016;Fu et al.,2013),已成为制约社会经济可持续发展的最主要因素,给我国农业生产和抗旱减灾工作带来前所未有的挑战和困难(吕娟等,2011;张强等,2014;Huang et al.,2016). ...
防旱抗旱减灾长效机制的构建策略探究
1
2020
... 农业防旱减灾是一项复杂的社会系统工程,涉及旱灾发生前的监测、预报、预警和防御,以及灾害发生时抗灾、灾后救灾和重建等多个方面.依照“工程措施与非工程措施相结合”的原则,在加强水利工程设施建设的同时,还需强化政策法规、监测预警、旱中保障等抗旱非工程措施,构建旱情监测预警和决策系统,提高抗旱减灾工作的科学化、现代化和规范化水平(姚国章和袁敏,2010;周广胜等,2016).鉴于旱灾对农业生产影响较重,急需建立一个科学有效的农业旱灾保障机制,但目前我国这种保障机制仍较滞后,相应的农业保险等项目发展缓慢(姚国章和袁敏,2010;周广胜等,2016).我国在干旱灾害管理方面的法律法规和政策尚有待健全,需借鉴欧美等发达国家经验,由被动的应急抗旱向主动的防旱模式转变(毛晓华,2020;屈艳萍等,2013).同时,我国在各类干旱(包括气象、农业、水文等)监测方面已开展了大量研究工作,但缺乏各部门间的有效协作机制,即气象、水文、农情、工业取水用水和相应供水部门之间的信息共享和联防联动.干旱整体评价指标构建上还需借鉴国外的干旱综合评价指标体系(康天军和李军波,2018;陈滢,2020).未来,我国还需制定更为详细的干旱灾害防御规划,让公众参与到干旱防灾、减灾工作中,构建全民参与的全社会防灾减灾新格局. ...
变化环境下我国干旱灾害演变趋势分析
8
2022
... 我国是一个农业大国,也是旱灾频发的国家,旱灾影响范围广、历时长、危害重.据统计,我国每年因干旱受灾的农田面积多达2 600万hm2,粮食减产高达50%~75%(姚玉璧等,2013;张强等,2014).近年来,受气候变化和水资源短缺的影响,我国农作物受旱、成灾和绝收面积不断扩大,在北方干旱形势依然严峻的情况下南方干旱出现明显增加和加重趋势,新旱区逐渐扩展,局部性、区域性干旱灾害频频出现,损失巨大(张强等,2014;康蕾和张红旗,2014;倪深海等,2022;张强,2022),严重威胁着国家的粮食安全和生态安全(王春乙,2007;张强等,2012a;张强等,2012b;肖国举和李裕,2012;康蕾和张红旗,2014;周广胜等,2016;Fu et al.,2013),已成为制约社会经济可持续发展的最主要因素,给我国农业生产和抗旱减灾工作带来前所未有的挑战和困难(吕娟等,2011;张强等,2014;Huang et al.,2016). ...
... 除了2022年重大农业干旱以外,回顾了1949—2020年的干旱状况,总体来说,全国发生轻旱、中旱、重旱以上的年份分别有55、46、26 a,发生频率分别为76.4%、63.9%、36.1%,发生特旱的年份有13 a,其频率为18.1%.多年来,因旱受灾和成灾面积分别为1 998.13、897.62万hm2,粮食损失1 630万t.其中,1949—2000年全国干旱受灾率、成灾率和因旱粮食损失率都呈现增加趋势,每10 a增速分别为1.7%、1.3%和0.6%,而2001—2020年干旱灾害呈减弱趋势.如果将第一个年代(1949—1959年)的成灾率和因旱粮食损失率作为基准看作1.0,则2000—2009年成灾率和损失率最高,分别是基准年代的3.9倍和2.9倍(图5),究其原因主要是2000年和2001年北方地区和华东、华中地区发生了特旱,因旱粮食损失较大,分别为5 996万t和5 480万t,相应的损失率分别达11.5%和10.8%,分别位列近70 a的第1位和第2位(倪深海等,2022). ...
... 1949—2020年不同年代我国粮食因旱受灾率、成灾率和损失率与1949—1959年比值变化(倪深海等,
2022)
Changes of the ratio of drought-affected rate and disaster-formative rate and loss rate due to drought in different decades in China during 1949-2020 compared with 1949-1959 (Ni et al., <xref ref-type="bibr" rid="b22">2022</xref>)Fig.5![]()
图6是1980年以来全国9大地区重旱以上灾害发生频率.可以看出,1980年以后我国北方的内蒙古、西北地区、东北地区和黄淮海地区干旱灾害发生频繁且较为严重,重旱以上发生频率分别为45.0%、40.0%、35.0%和25.0%,受灾率和成灾率均超过15.0%和10.0%(倪深海等,2022).从干旱发生范围来看,1980年以前发生重旱以上的区域波及10个省(市、区),而1980年以后扩增到16个省(市、区)(倪深海等,2022).受气候变暖影响,近50 a来西北地区干旱频率、强度和受灾面积增加,损失加重.其中,春旱、秋旱频次增加,夏旱频次减少,春旱、秋旱多于夏旱,特、重旱多出现在春季,主要发生在西北地区东部,尤其是甘肃河东地区,如1995年干旱最严重,受灾面积208.7万hm2,成灾面积170.8万hm2,成灾率为45%,粮食减产150万t;其次是2000年,成灾率达34.9%,粮食减产135万t(邓振镛等,2007).预计未来一段时间内,如果不采取有效应对措施,在气候变暖背景下到2030年中国种植业生产力总体可能下降5%~10%,其中小麦、水稻和玉米三大作物均以降低为主,2050年后受影响会更大(IPCC,2022). ...
... Changes of the ratio of drought-affected rate and disaster-formative rate and loss rate due to drought in different decades in China during 1949-2020 compared with 1949-1959 (Ni et al.,
2022)
Fig.5![]()
图6是1980年以来全国9大地区重旱以上灾害发生频率.可以看出,1980年以后我国北方的内蒙古、西北地区、东北地区和黄淮海地区干旱灾害发生频繁且较为严重,重旱以上发生频率分别为45.0%、40.0%、35.0%和25.0%,受灾率和成灾率均超过15.0%和10.0%(倪深海等,2022).从干旱发生范围来看,1980年以前发生重旱以上的区域波及10个省(市、区),而1980年以后扩增到16个省(市、区)(倪深海等,2022).受气候变暖影响,近50 a来西北地区干旱频率、强度和受灾面积增加,损失加重.其中,春旱、秋旱频次增加,夏旱频次减少,春旱、秋旱多于夏旱,特、重旱多出现在春季,主要发生在西北地区东部,尤其是甘肃河东地区,如1995年干旱最严重,受灾面积208.7万hm2,成灾面积170.8万hm2,成灾率为45%,粮食减产150万t;其次是2000年,成灾率达34.9%,粮食减产135万t(邓振镛等,2007).预计未来一段时间内,如果不采取有效应对措施,在气候变暖背景下到2030年中国种植业生产力总体可能下降5%~10%,其中小麦、水稻和玉米三大作物均以降低为主,2050年后受影响会更大(IPCC,2022). ...
... 图6是1980年以来全国9大地区重旱以上灾害发生频率.可以看出,1980年以后我国北方的内蒙古、西北地区、东北地区和黄淮海地区干旱灾害发生频繁且较为严重,重旱以上发生频率分别为45.0%、40.0%、35.0%和25.0%,受灾率和成灾率均超过15.0%和10.0%(倪深海等,2022).从干旱发生范围来看,1980年以前发生重旱以上的区域波及10个省(市、区),而1980年以后扩增到16个省(市、区)(倪深海等,2022).受气候变暖影响,近50 a来西北地区干旱频率、强度和受灾面积增加,损失加重.其中,春旱、秋旱频次增加,夏旱频次减少,春旱、秋旱多于夏旱,特、重旱多出现在春季,主要发生在西北地区东部,尤其是甘肃河东地区,如1995年干旱最严重,受灾面积208.7万hm2,成灾面积170.8万hm2,成灾率为45%,粮食减产150万t;其次是2000年,成灾率达34.9%,粮食减产135万t(邓振镛等,2007).预计未来一段时间内,如果不采取有效应对措施,在气候变暖背景下到2030年中国种植业生产力总体可能下降5%~10%,其中小麦、水稻和玉米三大作物均以降低为主,2050年后受影响会更大(IPCC,2022). ...
... ).从干旱发生范围来看,1980年以前发生重旱以上的区域波及10个省(市、区),而1980年以后扩增到16个省(市、区)(倪深海等,2022).受气候变暖影响,近50 a来西北地区干旱频率、强度和受灾面积增加,损失加重.其中,春旱、秋旱频次增加,夏旱频次减少,春旱、秋旱多于夏旱,特、重旱多出现在春季,主要发生在西北地区东部,尤其是甘肃河东地区,如1995年干旱最严重,受灾面积208.7万hm2,成灾面积170.8万hm2,成灾率为45%,粮食减产150万t;其次是2000年,成灾率达34.9%,粮食减产135万t(邓振镛等,2007).预计未来一段时间内,如果不采取有效应对措施,在气候变暖背景下到2030年中国种植业生产力总体可能下降5%~10%,其中小麦、水稻和玉米三大作物均以降低为主,2050年后受影响会更大(IPCC,2022). ...
... 1980—2020年全国重旱以上频率分布(倪深海等,
2022)
The frequency distribution of severe drought and above in China from 1980 to 2020 (Ni et al., <xref ref-type="bibr" rid="b22">2022</xref>)Fig.6![]() 2 干旱对粮食作物生产的影响机制回顾
2 干旱对粮食作物生产的影响机制回顾干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
... The frequency distribution of severe drought and above in China from 1980 to 2020 (Ni et al.,
2022)
Fig.6![]() 2 干旱对粮食作物生产的影响机制回顾
2 干旱对粮食作物生产的影响机制回顾干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
国际干旱灾害管理实践及对我国的启示
1
2013
... 农业防旱减灾是一项复杂的社会系统工程,涉及旱灾发生前的监测、预报、预警和防御,以及灾害发生时抗灾、灾后救灾和重建等多个方面.依照“工程措施与非工程措施相结合”的原则,在加强水利工程设施建设的同时,还需强化政策法规、监测预警、旱中保障等抗旱非工程措施,构建旱情监测预警和决策系统,提高抗旱减灾工作的科学化、现代化和规范化水平(姚国章和袁敏,2010;周广胜等,2016).鉴于旱灾对农业生产影响较重,急需建立一个科学有效的农业旱灾保障机制,但目前我国这种保障机制仍较滞后,相应的农业保险等项目发展缓慢(姚国章和袁敏,2010;周广胜等,2016).我国在干旱灾害管理方面的法律法规和政策尚有待健全,需借鉴欧美等发达国家经验,由被动的应急抗旱向主动的防旱模式转变(毛晓华,2020;屈艳萍等,2013).同时,我国在各类干旱(包括气象、农业、水文等)监测方面已开展了大量研究工作,但缺乏各部门间的有效协作机制,即气象、水文、农情、工业取水用水和相应供水部门之间的信息共享和联防联动.干旱整体评价指标构建上还需借鉴国外的干旱综合评价指标体系(康天军和李军波,2018;陈滢,2020).未来,我国还需制定更为详细的干旱灾害防御规划,让公众参与到干旱防灾、减灾工作中,构建全民参与的全社会防灾减灾新格局. ...
科学应对农业干旱
6
2011
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 第一,选择高产优质及抗旱、耐旱品种.培育与干旱相适应的作物新品种和优良品种,或者改良品种,特别是兼顾抗旱与水资源高效利用的品种(山仑,2011;张强等,2012b).开展水稻、玉米、小麦、马铃薯等作物新品种选育,对提高主粮生产能力具有重要作用.改良作物抗旱性的途径主要分为生物工程技术和生物激素鉴定两方面,在生物工程技术中传统育种利用自然等位基因适应特征的遗传多样性来改良植物抗旱性,基因组精准编辑工具和全基因组关联分析(Genome-wide Association Study,GWAS)等新技术的出现,在挖掘可提高抗旱性和产量的等位基因方面具有巨大应用潜力.生物刺激素中,一些小肽或激素激动剂的鉴定有助于精细调控干旱响应,在提高抗旱性同时保持了作物产量.这些技术的应用转化可为作物生产提供应对干旱的新策略(Gupta et al.,2020). ...
... 第三,在水资源缺乏的灌溉区和干旱山区推行水旱并举方略,逐步建立灌溉农业、旱地农业、设施农业和半旱地农业并存的农业用水新格局(山仑,2011),在黄淮海地区已有主动采用半旱地农业的实例,如山东恒台县采用的低定额灌溉条件下保持高产已初见成效,节水灌溉、少灌的半旱地轮作体系,也是比较成功的经验之一(山仑,2011).灌溉区调整灌溉模式,采用滴灌、喷灌等节水灌溉技术.在旱作农业区,根据农作物各生育期耗水亏损值进行分阶段、分次补充灌溉,在有条件的地区,基于物联网技术进行精细化灌溉和调控用水.开发农业用水新水源,如垄沟集雨、保墒集水、蓄集雨水、再生水、凝结水等都可作为补充水源供给农田,也可通过开发空中水资源补充陆地水资源不足(山仑,2011;张强等,2012b;肖国举和李裕,2012;王润元等,2015;熊友才和李凤民,2014). ...
... ),在黄淮海地区已有主动采用半旱地农业的实例,如山东恒台县采用的低定额灌溉条件下保持高产已初见成效,节水灌溉、少灌的半旱地轮作体系,也是比较成功的经验之一(山仑,2011).灌溉区调整灌溉模式,采用滴灌、喷灌等节水灌溉技术.在旱作农业区,根据农作物各生育期耗水亏损值进行分阶段、分次补充灌溉,在有条件的地区,基于物联网技术进行精细化灌溉和调控用水.开发农业用水新水源,如垄沟集雨、保墒集水、蓄集雨水、再生水、凝结水等都可作为补充水源供给农田,也可通过开发空中水资源补充陆地水资源不足(山仑,2011;张强等,2012b;肖国举和李裕,2012;王润元等,2015;熊友才和李凤民,2014). ...
... ).灌溉区调整灌溉模式,采用滴灌、喷灌等节水灌溉技术.在旱作农业区,根据农作物各生育期耗水亏损值进行分阶段、分次补充灌溉,在有条件的地区,基于物联网技术进行精细化灌溉和调控用水.开发农业用水新水源,如垄沟集雨、保墒集水、蓄集雨水、再生水、凝结水等都可作为补充水源供给农田,也可通过开发空中水资源补充陆地水资源不足(山仑,2011;张强等,2012b;肖国举和李裕,2012;王润元等,2015;熊友才和李凤民,2014). ...
... 第四,因地制宜推广保护性耕作措施,可同时起到保水、保土、培肥、增产的综合作用,如垄沟种植、垄沟径流集水、地膜覆盖、秸秆覆盖、免耕栽培等技术.采取多种形式的带状间作为中心的保护性耕作技术,缓解气候变暖加剧引起的水资源供求矛盾.推行麦类等条播作物与马铃薯、玉米、大豆等穴播作物或主粮作物与畜草合理间作、轮作等技术;合理套作,增加复种指数,提高耕地利用效率,如小麦套种马铃薯、小麦套种玉米等,可不同程度地提高作物产量,相当于在半干旱偏旱区的旱作农田上实现一年二熟(山仑,2011;张强等,2012a;张强等,2012b;肖国举和李裕,2012;鲍文中和周广胜,2017;Zhao et al.,2012;Zhao et al.,2014). ...
旱情评定与灾情指标之探讨
2
1994
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 在农业干旱监测中,农业干旱指标主要有气象指标、土壤墒情指标、作物生理生态指标和一些其他指标.气象指标包括降水距平百分数、连续无有效降水日数、湿度指标和Z指数等(孙荣强,1994;姚玉璧等,2013;国家气象中心等,2015;Zhang et al.,2009).土壤墒情指标包括土壤相对湿度、土壤有效水分存储量、土壤水分旱灾指数等(国家气象中心等,2015;Hollinger et al.,1993;Dai et al.,2004).作物生理生态指标主要是一些基于植株生长和光合特性的生理参数,用来衡量植株水分亏缺状况,如光合速率、蒸腾速率、气孔导度、复水后的光合恢复、叶水势、茎秆水势、叶绿素、叶片相对含水量、叶片扩展速率、作物水分胁迫指数、冠层温度、块茎碳同位素鉴别等(姚玉璧等,2013;张强等,2014;赵鸿等,2018;纪瑞鹏等,2019;Jefferies,1993;Medrano et al.,2002;Zakaluk and Ranjan,2006;Ricardo,2012;Ramirez et al.,2016),这些基于植株的监测指标更加方便、适时,但有些地区由于没有专用测量仪器而导致部分指标不能广泛应用. ...
我国主要农业气象灾害机理与监测研究进展
2
2005
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... ;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
黄土高原半干旱区马铃薯气候适宜度模拟及其时空变化特征
1
2017
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
4
2007
... 干旱灾害(简称“旱灾”),是指一段时间内降水量比常年明显偏少,造成某一地区经济社会活动(尤其是农业生产)和人类生活遭受较大危害的现象,普遍存在于世界各地,频繁发生于不同时期(Wilhite,2000;Li et al.,2015).旱灾是气象干旱、农业干旱、水文干旱、社会经济干旱和生态干旱之间的一种链状传递和转化过程,一个完整的干旱事件包括干旱历时、严重程度和平均强度(姚玉璧等,2013;张强等,2014;Heffernan,2013).全球每年因干旱导致的经济损失达60~80亿美元,2021年甚至高达121亿美元(American Meteorological Society, 1997;应急管理部-教育部减灾与应急管理研究院等,2022),远高于其他气象灾害造成的损失(张强等,2014;Wilhite,2000),受到高度关注(王春乙,2007;吕娟等,2011;Turner et al., 2011;Gupta et al.,2020). ...
... 农业干旱是农作物生长发育过程中,因降雨不足或长期无雨造成大气干旱、土壤缺水,致使作物得不到适时适量的水分补给,无法满足正常需水,影响其光合过程及生物量积累,最终造成农作物减产甚至绝收.农业干旱是影响农业生产的重要因素,也是农业生产面临的最大风险之一(王春乙,2007;肖国举和李裕,2012;张强等,2017;Mirzabaev et al.,2023).农业生产决定着粮食产量的稳定,是国家战略性、基础性核心产业,农作物种植是农业的“芯片”,是促进农业长期稳定发展,保障国家粮食安全的根本.干旱缺水直接影响农作物的生长发育和产量形成,造成粮食产量下降、品质降低,从而增加粮食生产供给的不稳定性和风险,直接威胁粮食安全,成为制约社会经济发展的重要因素之一(姚玉璧等,2013;张强等,2008;张强等,2020;IPCC,2007;IPCC,2022;Mirzabaev et al.,2023),长期以来农业旱灾对粮食安全的影响是学者和政府部门关注的重点和焦点.农业干旱对作物的影响从微观到宏观表现在细胞、生理和形态等不同层次水平上,其影响程度取决于干旱开始时间、发展和持续时间、严重程度及农作物种类和品种、所处生育期等因素(赵鸿等,2016;Ulrich et al.,2019;Gupta et al.,2020). ...
... 我国是一个农业大国,也是旱灾频发的国家,旱灾影响范围广、历时长、危害重.据统计,我国每年因干旱受灾的农田面积多达2 600万hm2,粮食减产高达50%~75%(姚玉璧等,2013;张强等,2014).近年来,受气候变化和水资源短缺的影响,我国农作物受旱、成灾和绝收面积不断扩大,在北方干旱形势依然严峻的情况下南方干旱出现明显增加和加重趋势,新旱区逐渐扩展,局部性、区域性干旱灾害频频出现,损失巨大(张强等,2014;康蕾和张红旗,2014;倪深海等,2022;张强,2022),严重威胁着国家的粮食安全和生态安全(王春乙,2007;张强等,2012a;张强等,2012b;肖国举和李裕,2012;康蕾和张红旗,2014;周广胜等,2016;Fu et al.,2013),已成为制约社会经济可持续发展的最主要因素,给我国农业生产和抗旱减灾工作带来前所未有的挑战和困难(吕娟等,2011;张强等,2014;Huang et al.,2016). ...
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
4
2015
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 第二,根据气候资源特点,调整种植结构和种植制度以趋利避害.在半干旱地区通过实行“压夏扩秋”(即压缩春小麦、春玉米等春季作物种植比例)措施,扩大适应干旱能力较强且与降雨同季的马铃薯、大豆等秋季作物种植面积,应对干旱的季节变化特征.在以雨养农业为主的陇东黄土高原区,农业生产中稳定冬小麦种植面积,增加玉米面积,发展马铃薯、豆类、糜子、谷子等抗旱性较强的作物(邓振镛等,2007;张强等,2012b;肖国举和李裕,2012).合理安排和调整作物种植面积和布局,加强水热资源的合理开发利用和管理,变被动抗旱为主动抗旱,管好、用好当地水资源,充分利用大气降水(张强等,2012b;王润元等,2015;鲍文中和周广胜,2017). ...
... 第三,在水资源缺乏的灌溉区和干旱山区推行水旱并举方略,逐步建立灌溉农业、旱地农业、设施农业和半旱地农业并存的农业用水新格局(山仑,2011),在黄淮海地区已有主动采用半旱地农业的实例,如山东恒台县采用的低定额灌溉条件下保持高产已初见成效,节水灌溉、少灌的半旱地轮作体系,也是比较成功的经验之一(山仑,2011).灌溉区调整灌溉模式,采用滴灌、喷灌等节水灌溉技术.在旱作农业区,根据农作物各生育期耗水亏损值进行分阶段、分次补充灌溉,在有条件的地区,基于物联网技术进行精细化灌溉和调控用水.开发农业用水新水源,如垄沟集雨、保墒集水、蓄集雨水、再生水、凝结水等都可作为补充水源供给农田,也可通过开发空中水资源补充陆地水资源不足(山仑,2011;张强等,2012b;肖国举和李裕,2012;王润元等,2015;熊友才和李凤民,2014). ...
... 第五,改进施肥方式和用量,测土施肥,配合深施、混施等施肥方式,提高肥效和作物对营养元素的利用率,如玉米施氮增效不仅可以获得较高产量、水分和氮素利用效率,还能降低硝态氮的残留量,有效减缓土壤污染(肖国举和李裕,2012;Qiang et al.,2019a).冬小麦测土施肥,可用于诊断其氮营养状况,提高水分利用效率、增加叶面积指数和干物质积累(肖国举和李裕,2012;Qiang et al.,2019b).未来半干旱地区的小麦生产,可视温度、CO2浓度升高情况适量采用低磷肥、低氮肥投入,这将更经济、适宜(王润元等,2015;肖国举和李裕,2012;鲍文中和周广胜,2017). ...
国际社会如何应对干旱灾害
1
2011
... 以色列国家面对水资源严重贫乏的现状,首先制定了《水法》,颁布全国水资源管理和用水政策,实施工农业用水配额,大力推行农业节水灌溉、废水处理灌溉、微咸水灌溉等措施,将每一滴水都用到了极致(韩萍,2002).瑞典、荷兰、德国等欧洲国家利用人工补给含水层来增加水资源量,极力发挥地下水库调节作用以抵御干旱风险(https://www.docin.com/p-2012740929.html).欧盟也制定了一系列旱灾风险管理战略,从水资源危机管理向干旱风险管理转变(https://world.qianlong.com/2022/0830/7575142.shtml).埃塞俄比亚、肯尼亚、索马里等非洲国家则通过积极引进抗旱作物和抗旱技术应对农业干旱造成的粮食短缺问题(吴爱民,2011),如为缓解干旱造成的粮食短缺,引进了具有耐旱性的玉米新品种,水稻高杆品种变为矮杆,并辅配农药和农业机械,解决了19个发展中国家的粮食自给问题(https://world.qianlong.com/2022/0830/7575142.shtml). ...
水文干旱对气象干旱的响应及其临界条件
1
2017
... 农业干旱应对主要包括农业干旱的减轻和防御两方面,农业干旱的发生、发展与气象干旱、水文干旱等其他类型干旱紧密相关(吴杰峰等,2017),因此在应对和防御农业干旱时,不仅要关注农业干旱,还要加强其他类型干旱的监测和管理.农业干旱的减轻措施是指用来抵御干旱灾害的不利影响、环境恶化以及技术风险的工程措施(如基础设施、修筑堤坝、工程建设等)或非工程措施(如政策法规、政府决策、公众意识等).农业干旱的防御措施是指在干旱灾害发生前制定的政策规划、制度以及灾害的监测、预测或预警,以确保灾害发生时各方面的协调与有效响应(冯定原和邱新法,1995;陆亚龙和肖功建,2001).粮食生产对干旱的减轻、防御和适应技术对策见图8. ...
9
2012
... 农业干旱是农作物生长发育过程中,因降雨不足或长期无雨造成大气干旱、土壤缺水,致使作物得不到适时适量的水分补给,无法满足正常需水,影响其光合过程及生物量积累,最终造成农作物减产甚至绝收.农业干旱是影响农业生产的重要因素,也是农业生产面临的最大风险之一(王春乙,2007;肖国举和李裕,2012;张强等,2017;Mirzabaev et al.,2023).农业生产决定着粮食产量的稳定,是国家战略性、基础性核心产业,农作物种植是农业的“芯片”,是促进农业长期稳定发展,保障国家粮食安全的根本.干旱缺水直接影响农作物的生长发育和产量形成,造成粮食产量下降、品质降低,从而增加粮食生产供给的不稳定性和风险,直接威胁粮食安全,成为制约社会经济发展的重要因素之一(姚玉璧等,2013;张强等,2008;张强等,2020;IPCC,2007;IPCC,2022;Mirzabaev et al.,2023),长期以来农业旱灾对粮食安全的影响是学者和政府部门关注的重点和焦点.农业干旱对作物的影响从微观到宏观表现在细胞、生理和形态等不同层次水平上,其影响程度取决于干旱开始时间、发展和持续时间、严重程度及农作物种类和品种、所处生育期等因素(赵鸿等,2016;Ulrich et al.,2019;Gupta et al.,2020). ...
... 我国是一个农业大国,也是旱灾频发的国家,旱灾影响范围广、历时长、危害重.据统计,我国每年因干旱受灾的农田面积多达2 600万hm2,粮食减产高达50%~75%(姚玉璧等,2013;张强等,2014).近年来,受气候变化和水资源短缺的影响,我国农作物受旱、成灾和绝收面积不断扩大,在北方干旱形势依然严峻的情况下南方干旱出现明显增加和加重趋势,新旱区逐渐扩展,局部性、区域性干旱灾害频频出现,损失巨大(张强等,2014;康蕾和张红旗,2014;倪深海等,2022;张强,2022),严重威胁着国家的粮食安全和生态安全(王春乙,2007;张强等,2012a;张强等,2012b;肖国举和李裕,2012;康蕾和张红旗,2014;周广胜等,2016;Fu et al.,2013),已成为制约社会经济可持续发展的最主要因素,给我国农业生产和抗旱减灾工作带来前所未有的挑战和困难(吕娟等,2011;张强等,2014;Huang et al.,2016). ...
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 第二,根据气候资源特点,调整种植结构和种植制度以趋利避害.在半干旱地区通过实行“压夏扩秋”(即压缩春小麦、春玉米等春季作物种植比例)措施,扩大适应干旱能力较强且与降雨同季的马铃薯、大豆等秋季作物种植面积,应对干旱的季节变化特征.在以雨养农业为主的陇东黄土高原区,农业生产中稳定冬小麦种植面积,增加玉米面积,发展马铃薯、豆类、糜子、谷子等抗旱性较强的作物(邓振镛等,2007;张强等,2012b;肖国举和李裕,2012).合理安排和调整作物种植面积和布局,加强水热资源的合理开发利用和管理,变被动抗旱为主动抗旱,管好、用好当地水资源,充分利用大气降水(张强等,2012b;王润元等,2015;鲍文中和周广胜,2017). ...
... 第三,在水资源缺乏的灌溉区和干旱山区推行水旱并举方略,逐步建立灌溉农业、旱地农业、设施农业和半旱地农业并存的农业用水新格局(山仑,2011),在黄淮海地区已有主动采用半旱地农业的实例,如山东恒台县采用的低定额灌溉条件下保持高产已初见成效,节水灌溉、少灌的半旱地轮作体系,也是比较成功的经验之一(山仑,2011).灌溉区调整灌溉模式,采用滴灌、喷灌等节水灌溉技术.在旱作农业区,根据农作物各生育期耗水亏损值进行分阶段、分次补充灌溉,在有条件的地区,基于物联网技术进行精细化灌溉和调控用水.开发农业用水新水源,如垄沟集雨、保墒集水、蓄集雨水、再生水、凝结水等都可作为补充水源供给农田,也可通过开发空中水资源补充陆地水资源不足(山仑,2011;张强等,2012b;肖国举和李裕,2012;王润元等,2015;熊友才和李凤民,2014). ...
... 第四,因地制宜推广保护性耕作措施,可同时起到保水、保土、培肥、增产的综合作用,如垄沟种植、垄沟径流集水、地膜覆盖、秸秆覆盖、免耕栽培等技术.采取多种形式的带状间作为中心的保护性耕作技术,缓解气候变暖加剧引起的水资源供求矛盾.推行麦类等条播作物与马铃薯、玉米、大豆等穴播作物或主粮作物与畜草合理间作、轮作等技术;合理套作,增加复种指数,提高耕地利用效率,如小麦套种马铃薯、小麦套种玉米等,可不同程度地提高作物产量,相当于在半干旱偏旱区的旱作农田上实现一年二熟(山仑,2011;张强等,2012a;张强等,2012b;肖国举和李裕,2012;鲍文中和周广胜,2017;Zhao et al.,2012;Zhao et al.,2014). ...
... 第五,改进施肥方式和用量,测土施肥,配合深施、混施等施肥方式,提高肥效和作物对营养元素的利用率,如玉米施氮增效不仅可以获得较高产量、水分和氮素利用效率,还能降低硝态氮的残留量,有效减缓土壤污染(肖国举和李裕,2012;Qiang et al.,2019a).冬小麦测土施肥,可用于诊断其氮营养状况,提高水分利用效率、增加叶面积指数和干物质积累(肖国举和李裕,2012;Qiang et al.,2019b).未来半干旱地区的小麦生产,可视温度、CO2浓度升高情况适量采用低磷肥、低氮肥投入,这将更经济、适宜(王润元等,2015;肖国举和李裕,2012;鲍文中和周广胜,2017). ...
... ).冬小麦测土施肥,可用于诊断其氮营养状况,提高水分利用效率、增加叶面积指数和干物质积累(肖国举和李裕,2012;Qiang et al.,2019b).未来半干旱地区的小麦生产,可视温度、CO2浓度升高情况适量采用低磷肥、低氮肥投入,这将更经济、适宜(王润元等,2015;肖国举和李裕,2012;鲍文中和周广胜,2017). ...
... ;肖国举和李裕,2012;鲍文中和周广胜,2017). ...
2
2014
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 第三,在水资源缺乏的灌溉区和干旱山区推行水旱并举方略,逐步建立灌溉农业、旱地农业、设施农业和半旱地农业并存的农业用水新格局(山仑,2011),在黄淮海地区已有主动采用半旱地农业的实例,如山东恒台县采用的低定额灌溉条件下保持高产已初见成效,节水灌溉、少灌的半旱地轮作体系,也是比较成功的经验之一(山仑,2011).灌溉区调整灌溉模式,采用滴灌、喷灌等节水灌溉技术.在旱作农业区,根据农作物各生育期耗水亏损值进行分阶段、分次补充灌溉,在有条件的地区,基于物联网技术进行精细化灌溉和调控用水.开发农业用水新水源,如垄沟集雨、保墒集水、蓄集雨水、再生水、凝结水等都可作为补充水源供给农田,也可通过开发空中水资源补充陆地水资源不足(山仑,2011;张强等,2012b;肖国举和李裕,2012;王润元等,2015;熊友才和李凤民,2014). ...
干旱预警系统建设的国际经验与借鉴
3
2010
... 第六,加强农业干旱监测预测预警,强化防旱减灾应对准备.在出现旱情的重点地区加强土壤墒情监测和定点调查,构建区域信息共享平台,及时掌握旱情发展蔓延动态;加快构建农业农村气象灾害预警信息发布系统,搭建多种渠道的气象服务信息传播平台,建立气象灾害风险评估体系,研发基于影响的农业干旱监测预测预警技术,针对农业生产中重要环节和农作物生长发育的水分需求,提升农业防旱减灾服务能力,定期发布土壤墒情监测公报、干旱监测公报、农业气象旬(周)报等农业气象服务产品(姚国章和袁敏,2010;张强等,2014;周广胜等,2016).总之,在确保国家和区域(或省、市、区)粮、棉、油需求安全前提下,充分利用水热资源优势,压缩高耗水作物和品种的种植面积,实行农业补贴政策,实现农业经济和水资源安全协调发展(鲍文中和周广胜,2017). ...
... 农业防旱减灾是一项复杂的社会系统工程,涉及旱灾发生前的监测、预报、预警和防御,以及灾害发生时抗灾、灾后救灾和重建等多个方面.依照“工程措施与非工程措施相结合”的原则,在加强水利工程设施建设的同时,还需强化政策法规、监测预警、旱中保障等抗旱非工程措施,构建旱情监测预警和决策系统,提高抗旱减灾工作的科学化、现代化和规范化水平(姚国章和袁敏,2010;周广胜等,2016).鉴于旱灾对农业生产影响较重,急需建立一个科学有效的农业旱灾保障机制,但目前我国这种保障机制仍较滞后,相应的农业保险等项目发展缓慢(姚国章和袁敏,2010;周广胜等,2016).我国在干旱灾害管理方面的法律法规和政策尚有待健全,需借鉴欧美等发达国家经验,由被动的应急抗旱向主动的防旱模式转变(毛晓华,2020;屈艳萍等,2013).同时,我国在各类干旱(包括气象、农业、水文等)监测方面已开展了大量研究工作,但缺乏各部门间的有效协作机制,即气象、水文、农情、工业取水用水和相应供水部门之间的信息共享和联防联动.干旱整体评价指标构建上还需借鉴国外的干旱综合评价指标体系(康天军和李军波,2018;陈滢,2020).未来,我国还需制定更为详细的干旱灾害防御规划,让公众参与到干旱防灾、减灾工作中,构建全民参与的全社会防灾减灾新格局. ...
... ).鉴于旱灾对农业生产影响较重,急需建立一个科学有效的农业旱灾保障机制,但目前我国这种保障机制仍较滞后,相应的农业保险等项目发展缓慢(姚国章和袁敏,2010;周广胜等,2016).我国在干旱灾害管理方面的法律法规和政策尚有待健全,需借鉴欧美等发达国家经验,由被动的应急抗旱向主动的防旱模式转变(毛晓华,2020;屈艳萍等,2013).同时,我国在各类干旱(包括气象、农业、水文等)监测方面已开展了大量研究工作,但缺乏各部门间的有效协作机制,即气象、水文、农情、工业取水用水和相应供水部门之间的信息共享和联防联动.干旱整体评价指标构建上还需借鉴国外的干旱综合评价指标体系(康天军和李军波,2018;陈滢,2020).未来,我国还需制定更为详细的干旱灾害防御规划,让公众参与到干旱防灾、减灾工作中,构建全民参与的全社会防灾减灾新格局. ...
干旱灾害风险评估技术及其科学问题与展望
6
2013
... 干旱灾害(简称“旱灾”),是指一段时间内降水量比常年明显偏少,造成某一地区经济社会活动(尤其是农业生产)和人类生活遭受较大危害的现象,普遍存在于世界各地,频繁发生于不同时期(Wilhite,2000;Li et al.,2015).旱灾是气象干旱、农业干旱、水文干旱、社会经济干旱和生态干旱之间的一种链状传递和转化过程,一个完整的干旱事件包括干旱历时、严重程度和平均强度(姚玉璧等,2013;张强等,2014;Heffernan,2013).全球每年因干旱导致的经济损失达60~80亿美元,2021年甚至高达121亿美元(American Meteorological Society, 1997;应急管理部-教育部减灾与应急管理研究院等,2022),远高于其他气象灾害造成的损失(张强等,2014;Wilhite,2000),受到高度关注(王春乙,2007;吕娟等,2011;Turner et al., 2011;Gupta et al.,2020). ...
... 农业干旱是农作物生长发育过程中,因降雨不足或长期无雨造成大气干旱、土壤缺水,致使作物得不到适时适量的水分补给,无法满足正常需水,影响其光合过程及生物量积累,最终造成农作物减产甚至绝收.农业干旱是影响农业生产的重要因素,也是农业生产面临的最大风险之一(王春乙,2007;肖国举和李裕,2012;张强等,2017;Mirzabaev et al.,2023).农业生产决定着粮食产量的稳定,是国家战略性、基础性核心产业,农作物种植是农业的“芯片”,是促进农业长期稳定发展,保障国家粮食安全的根本.干旱缺水直接影响农作物的生长发育和产量形成,造成粮食产量下降、品质降低,从而增加粮食生产供给的不稳定性和风险,直接威胁粮食安全,成为制约社会经济发展的重要因素之一(姚玉璧等,2013;张强等,2008;张强等,2020;IPCC,2007;IPCC,2022;Mirzabaev et al.,2023),长期以来农业旱灾对粮食安全的影响是学者和政府部门关注的重点和焦点.农业干旱对作物的影响从微观到宏观表现在细胞、生理和形态等不同层次水平上,其影响程度取决于干旱开始时间、发展和持续时间、严重程度及农作物种类和品种、所处生育期等因素(赵鸿等,2016;Ulrich et al.,2019;Gupta et al.,2020). ...
... 我国是一个农业大国,也是旱灾频发的国家,旱灾影响范围广、历时长、危害重.据统计,我国每年因干旱受灾的农田面积多达2 600万hm2,粮食减产高达50%~75%(姚玉璧等,2013;张强等,2014).近年来,受气候变化和水资源短缺的影响,我国农作物受旱、成灾和绝收面积不断扩大,在北方干旱形势依然严峻的情况下南方干旱出现明显增加和加重趋势,新旱区逐渐扩展,局部性、区域性干旱灾害频频出现,损失巨大(张强等,2014;康蕾和张红旗,2014;倪深海等,2022;张强,2022),严重威胁着国家的粮食安全和生态安全(王春乙,2007;张强等,2012a;张强等,2012b;肖国举和李裕,2012;康蕾和张红旗,2014;周广胜等,2016;Fu et al.,2013),已成为制约社会经济可持续发展的最主要因素,给我国农业生产和抗旱减灾工作带来前所未有的挑战和困难(吕娟等,2011;张强等,2014;Huang et al.,2016). ...
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 在农业干旱监测中,农业干旱指标主要有气象指标、土壤墒情指标、作物生理生态指标和一些其他指标.气象指标包括降水距平百分数、连续无有效降水日数、湿度指标和Z指数等(孙荣强,1994;姚玉璧等,2013;国家气象中心等,2015;Zhang et al.,2009).土壤墒情指标包括土壤相对湿度、土壤有效水分存储量、土壤水分旱灾指数等(国家气象中心等,2015;Hollinger et al.,1993;Dai et al.,2004).作物生理生态指标主要是一些基于植株生长和光合特性的生理参数,用来衡量植株水分亏缺状况,如光合速率、蒸腾速率、气孔导度、复水后的光合恢复、叶水势、茎秆水势、叶绿素、叶片相对含水量、叶片扩展速率、作物水分胁迫指数、冠层温度、块茎碳同位素鉴别等(姚玉璧等,2013;张强等,2014;赵鸿等,2018;纪瑞鹏等,2019;Jefferies,1993;Medrano et al.,2002;Zakaluk and Ranjan,2006;Ricardo,2012;Ramirez et al.,2016),这些基于植株的监测指标更加方便、适时,但有些地区由于没有专用测量仪器而导致部分指标不能广泛应用. ...
... ).作物生理生态指标主要是一些基于植株生长和光合特性的生理参数,用来衡量植株水分亏缺状况,如光合速率、蒸腾速率、气孔导度、复水后的光合恢复、叶水势、茎秆水势、叶绿素、叶片相对含水量、叶片扩展速率、作物水分胁迫指数、冠层温度、块茎碳同位素鉴别等(姚玉璧等,2013;张强等,2014;赵鸿等,2018;纪瑞鹏等,2019;Jefferies,1993;Medrano et al.,2002;Zakaluk and Ranjan,2006;Ricardo,2012;Ramirez et al.,2016),这些基于植株的监测指标更加方便、适时,但有些地区由于没有专用测量仪器而导致部分指标不能广泛应用. ...
1
2022
... 干旱灾害(简称“旱灾”),是指一段时间内降水量比常年明显偏少,造成某一地区经济社会活动(尤其是农业生产)和人类生活遭受较大危害的现象,普遍存在于世界各地,频繁发生于不同时期(Wilhite,2000;Li et al.,2015).旱灾是气象干旱、农业干旱、水文干旱、社会经济干旱和生态干旱之间的一种链状传递和转化过程,一个完整的干旱事件包括干旱历时、严重程度和平均强度(姚玉璧等,2013;张强等,2014;Heffernan,2013).全球每年因干旱导致的经济损失达60~80亿美元,2021年甚至高达121亿美元(American Meteorological Society, 1997;应急管理部-教育部减灾与应急管理研究院等,2022),远高于其他气象灾害造成的损失(张强等,2014;Wilhite,2000),受到高度关注(王春乙,2007;吕娟等,2011;Turner et al., 2011;Gupta et al.,2020). ...
干旱对小麦孕穗期叶片生理特性及产量的影响
1
2014
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
科学解读“2022年长江流域重大干旱”
3
2022
... 受全球气候变化和经济快速发展的影响,全球干旱呈现多发、加重趋势,中亚地区65%的区域表现为干旱程度加剧、强度持续增强,2022年全球极端干旱问题尤为突出(李稚等,2022;张强,2022;Metz et al.,2007;Li et al.,2015;IPCC,2022).随着全球气候变暖,水循环加快,陆地蒸散发和储水量减少,干旱半干旱区不断扩张,干旱持续加剧,干旱风险进一步加大,且存在较大的区域差异(张强等,2017;Salinger et al.,2005;Cook et al.,2007;Metz et al.,2007;Huang et al.,2016).联合国《2022年全球干旱数字》报告指出,自2000年以来全球干旱次数增加了29%,全球处于高温和干旱管理的“十字路口”(UNCCD, 2022).预计到2100年,极端干旱地区的干旱次数和强度将增加1%~30%(IPCC,2007). ...
... 我国是一个农业大国,也是旱灾频发的国家,旱灾影响范围广、历时长、危害重.据统计,我国每年因干旱受灾的农田面积多达2 600万hm2,粮食减产高达50%~75%(姚玉璧等,2013;张强等,2014).近年来,受气候变化和水资源短缺的影响,我国农作物受旱、成灾和绝收面积不断扩大,在北方干旱形势依然严峻的情况下南方干旱出现明显增加和加重趋势,新旱区逐渐扩展,局部性、区域性干旱灾害频频出现,损失巨大(张强等,2014;康蕾和张红旗,2014;倪深海等,2022;张强,2022),严重威胁着国家的粮食安全和生态安全(王春乙,2007;张强等,2012a;张强等,2012b;肖国举和李裕,2012;康蕾和张红旗,2014;周广胜等,2016;Fu et al.,2013),已成为制约社会经济可持续发展的最主要因素,给我国农业生产和抗旱减灾工作带来前所未有的挑战和困难(吕娟等,2011;张强等,2014;Huang et al.,2016). ...
... 我国旱灾呈多发、频发特征,农业受灾严重.2022年,我国西南、北方、长江中下游等多地区先后出现大范围、不同程度的严重干旱,覆盖河北、山西、山东、河南、安徽、湖北、陕西、甘肃和新疆等12个省(区),干旱持续日数长,长江流域的干旱时间超过50 d,受干旱影响的作物面积有387万hm2,其中93万hm2受干旱影响严重.据统计,7月以来,长江流域中旱及以上等级日数为45.6 d,较常年同期偏多31.0 d,为1961年以来历史同期最多;中旱及以上等级站数涉及667个,占全流域的96%(https://www.163.com/dy/article/HIEPM4LJ0514A42S.html),影响范围为1961年以来同期最大,严重干旱造成200多万 hm2 农田受灾,数亿人生活受到影响(张强,2022).据国家气候中心气象干旱监测显示,8月21日江苏南部、安徽南部、河南西南部、湖北中西部、浙江大部、福建大部、江西、湖南、贵州大部、重庆、四川大部、甘肃东南部地区和藏区中北部地区仍有轻、中度到中、重度气象干旱,部分地区有特旱(图4)(https://www.jnw.cc/yqzt/202208/1626135.html).进入9月,长江中下游地区仍持续干旱少雨,大部地区遭受夏秋连旱.9月28日,长江流域中下游大部地区仍存在中度及以上等级气象干旱,其中浙江西南部、安徽南部、湖北东部、湖南中南部、江西大部、福建大部等地有特旱(https://www.wuhan.com/xinwen/108763.html),对农业生产造成较大影响,大部分农作物产量有不同程度减产. ...
气候变化与西北地区粮食和食品安全
3
2012a
... 我国是一个农业大国,也是旱灾频发的国家,旱灾影响范围广、历时长、危害重.据统计,我国每年因干旱受灾的农田面积多达2 600万hm2,粮食减产高达50%~75%(姚玉璧等,2013;张强等,2014).近年来,受气候变化和水资源短缺的影响,我国农作物受旱、成灾和绝收面积不断扩大,在北方干旱形势依然严峻的情况下南方干旱出现明显增加和加重趋势,新旱区逐渐扩展,局部性、区域性干旱灾害频频出现,损失巨大(张强等,2014;康蕾和张红旗,2014;倪深海等,2022;张强,2022),严重威胁着国家的粮食安全和生态安全(王春乙,2007;张强等,2012a;张强等,2012b;肖国举和李裕,2012;康蕾和张红旗,2014;周广胜等,2016;Fu et al.,2013),已成为制约社会经济可持续发展的最主要因素,给我国农业生产和抗旱减灾工作带来前所未有的挑战和困难(吕娟等,2011;张强等,2014;Huang et al.,2016). ...
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 第四,因地制宜推广保护性耕作措施,可同时起到保水、保土、培肥、增产的综合作用,如垄沟种植、垄沟径流集水、地膜覆盖、秸秆覆盖、免耕栽培等技术.采取多种形式的带状间作为中心的保护性耕作技术,缓解气候变暖加剧引起的水资源供求矛盾.推行麦类等条播作物与马铃薯、玉米、大豆等穴播作物或主粮作物与畜草合理间作、轮作等技术;合理套作,增加复种指数,提高耕地利用效率,如小麦套种马铃薯、小麦套种玉米等,可不同程度地提高作物产量,相当于在半干旱偏旱区的旱作农田上实现一年二熟(山仑,2011;张强等,2012a;张强等,2012b;肖国举和李裕,2012;鲍文中和周广胜,2017;Zhao et al.,2012;Zhao et al.,2014). ...
全球气候变化对我国西北地区农业的影响
2
2008
... 农业干旱是农作物生长发育过程中,因降雨不足或长期无雨造成大气干旱、土壤缺水,致使作物得不到适时适量的水分补给,无法满足正常需水,影响其光合过程及生物量积累,最终造成农作物减产甚至绝收.农业干旱是影响农业生产的重要因素,也是农业生产面临的最大风险之一(王春乙,2007;肖国举和李裕,2012;张强等,2017;Mirzabaev et al.,2023).农业生产决定着粮食产量的稳定,是国家战略性、基础性核心产业,农作物种植是农业的“芯片”,是促进农业长期稳定发展,保障国家粮食安全的根本.干旱缺水直接影响农作物的生长发育和产量形成,造成粮食产量下降、品质降低,从而增加粮食生产供给的不稳定性和风险,直接威胁粮食安全,成为制约社会经济发展的重要因素之一(姚玉璧等,2013;张强等,2008;张强等,2020;IPCC,2007;IPCC,2022;Mirzabaev et al.,2023),长期以来农业旱灾对粮食安全的影响是学者和政府部门关注的重点和焦点.农业干旱对作物的影响从微观到宏观表现在细胞、生理和形态等不同层次水平上,其影响程度取决于干旱开始时间、发展和持续时间、严重程度及农作物种类和品种、所处生育期等因素(赵鸿等,2016;Ulrich et al.,2019;Gupta et al.,2020). ...
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
论气候变暖背景下干旱和干旱灾害风险特征与管理策略
8
2014
... 干旱灾害(简称“旱灾”),是指一段时间内降水量比常年明显偏少,造成某一地区经济社会活动(尤其是农业生产)和人类生活遭受较大危害的现象,普遍存在于世界各地,频繁发生于不同时期(Wilhite,2000;Li et al.,2015).旱灾是气象干旱、农业干旱、水文干旱、社会经济干旱和生态干旱之间的一种链状传递和转化过程,一个完整的干旱事件包括干旱历时、严重程度和平均强度(姚玉璧等,2013;张强等,2014;Heffernan,2013).全球每年因干旱导致的经济损失达60~80亿美元,2021年甚至高达121亿美元(American Meteorological Society, 1997;应急管理部-教育部减灾与应急管理研究院等,2022),远高于其他气象灾害造成的损失(张强等,2014;Wilhite,2000),受到高度关注(王春乙,2007;吕娟等,2011;Turner et al., 2011;Gupta et al.,2020). ...
... ),远高于其他气象灾害造成的损失(张强等,2014;Wilhite,2000),受到高度关注(王春乙,2007;吕娟等,2011;Turner et al., 2011;Gupta et al.,2020). ...
... 我国是一个农业大国,也是旱灾频发的国家,旱灾影响范围广、历时长、危害重.据统计,我国每年因干旱受灾的农田面积多达2 600万hm2,粮食减产高达50%~75%(姚玉璧等,2013;张强等,2014).近年来,受气候变化和水资源短缺的影响,我国农作物受旱、成灾和绝收面积不断扩大,在北方干旱形势依然严峻的情况下南方干旱出现明显增加和加重趋势,新旱区逐渐扩展,局部性、区域性干旱灾害频频出现,损失巨大(张强等,2014;康蕾和张红旗,2014;倪深海等,2022;张强,2022),严重威胁着国家的粮食安全和生态安全(王春乙,2007;张强等,2012a;张强等,2012b;肖国举和李裕,2012;康蕾和张红旗,2014;周广胜等,2016;Fu et al.,2013),已成为制约社会经济可持续发展的最主要因素,给我国农业生产和抗旱减灾工作带来前所未有的挑战和困难(吕娟等,2011;张强等,2014;Huang et al.,2016). ...
... ).近年来,受气候变化和水资源短缺的影响,我国农作物受旱、成灾和绝收面积不断扩大,在北方干旱形势依然严峻的情况下南方干旱出现明显增加和加重趋势,新旱区逐渐扩展,局部性、区域性干旱灾害频频出现,损失巨大(张强等,2014;康蕾和张红旗,2014;倪深海等,2022;张强,2022),严重威胁着国家的粮食安全和生态安全(王春乙,2007;张强等,2012a;张强等,2012b;肖国举和李裕,2012;康蕾和张红旗,2014;周广胜等,2016;Fu et al.,2013),已成为制约社会经济可持续发展的最主要因素,给我国农业生产和抗旱减灾工作带来前所未有的挑战和困难(吕娟等,2011;张强等,2014;Huang et al.,2016). ...
... ;张强等,2014;Huang et al.,2016). ...
... 2012年夏季美国遭遇了56 a来最严重旱灾,导致严重的粮食产量下降、品质降低和价格猛涨,其中玉米和小麦价格分别上涨60%和26%,引发了全球性的粮食危机.同年,受干旱气候影响,俄罗斯、乌克兰、中国等国家小麦、玉米主产区减产,也引起了不同程度的粮食减产和价格上涨(张强等,2014).2010年夏季俄罗斯遭受了130 a来最严重的干旱,导致近1/3的农作物受灾,因粮食减产而暂停小麦、玉米等农产品出口(https://www.docin.com/p-1435663470.html). 2016年越南南部遭遇近百年来的严重干旱,干旱对水稻影响尤为严重,湄公河三角洲区域农业受灾严重(张强等,2017).预计到21世纪末,气候变化将对非洲和中南美洲谷物产生负面影响,叠加物候影响,这将势必改变当前全球作物适宜种植区(https://m.thepaper.cn/baijiahao_20016300).2022年11月15日,联合国秘书长在二十国集团峰会上表示,如果不采取协调行动,今年的“粮食负担能力危机”可能在2023年变成严重的全球粮食短缺问题,并呼吁现在就共同行动起来,防止明年发生“严重的粮食灾难”(https://news.un.org/zh/story/2022/11/1112462). ...
... 在农业干旱监测中,农业干旱指标主要有气象指标、土壤墒情指标、作物生理生态指标和一些其他指标.气象指标包括降水距平百分数、连续无有效降水日数、湿度指标和Z指数等(孙荣强,1994;姚玉璧等,2013;国家气象中心等,2015;Zhang et al.,2009).土壤墒情指标包括土壤相对湿度、土壤有效水分存储量、土壤水分旱灾指数等(国家气象中心等,2015;Hollinger et al.,1993;Dai et al.,2004).作物生理生态指标主要是一些基于植株生长和光合特性的生理参数,用来衡量植株水分亏缺状况,如光合速率、蒸腾速率、气孔导度、复水后的光合恢复、叶水势、茎秆水势、叶绿素、叶片相对含水量、叶片扩展速率、作物水分胁迫指数、冠层温度、块茎碳同位素鉴别等(姚玉璧等,2013;张强等,2014;赵鸿等,2018;纪瑞鹏等,2019;Jefferies,1993;Medrano et al.,2002;Zakaluk and Ranjan,2006;Ricardo,2012;Ramirez et al.,2016),这些基于植株的监测指标更加方便、适时,但有些地区由于没有专用测量仪器而导致部分指标不能广泛应用. ...
... 第六,加强农业干旱监测预测预警,强化防旱减灾应对准备.在出现旱情的重点地区加强土壤墒情监测和定点调查,构建区域信息共享平台,及时掌握旱情发展蔓延动态;加快构建农业农村气象灾害预警信息发布系统,搭建多种渠道的气象服务信息传播平台,建立气象灾害风险评估体系,研发基于影响的农业干旱监测预测预警技术,针对农业生产中重要环节和农作物生长发育的水分需求,提升农业防旱减灾服务能力,定期发布土壤墒情监测公报、干旱监测公报、农业气象旬(周)报等农业气象服务产品(姚国章和袁敏,2010;张强等,2014;周广胜等,2016).总之,在确保国家和区域(或省、市、区)粮、棉、油需求安全前提下,充分利用水热资源优势,压缩高耗水作物和品种的种植面积,实行农业补贴政策,实现农业经济和水资源安全协调发展(鲍文中和周广胜,2017). ...
7
2012b
... 我国是一个农业大国,也是旱灾频发的国家,旱灾影响范围广、历时长、危害重.据统计,我国每年因干旱受灾的农田面积多达2 600万hm2,粮食减产高达50%~75%(姚玉璧等,2013;张强等,2014).近年来,受气候变化和水资源短缺的影响,我国农作物受旱、成灾和绝收面积不断扩大,在北方干旱形势依然严峻的情况下南方干旱出现明显增加和加重趋势,新旱区逐渐扩展,局部性、区域性干旱灾害频频出现,损失巨大(张强等,2014;康蕾和张红旗,2014;倪深海等,2022;张强,2022),严重威胁着国家的粮食安全和生态安全(王春乙,2007;张强等,2012a;张强等,2012b;肖国举和李裕,2012;康蕾和张红旗,2014;周广胜等,2016;Fu et al.,2013),已成为制约社会经济可持续发展的最主要因素,给我国农业生产和抗旱减灾工作带来前所未有的挑战和困难(吕娟等,2011;张强等,2014;Huang et al.,2016). ...
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 第一,选择高产优质及抗旱、耐旱品种.培育与干旱相适应的作物新品种和优良品种,或者改良品种,特别是兼顾抗旱与水资源高效利用的品种(山仑,2011;张强等,2012b).开展水稻、玉米、小麦、马铃薯等作物新品种选育,对提高主粮生产能力具有重要作用.改良作物抗旱性的途径主要分为生物工程技术和生物激素鉴定两方面,在生物工程技术中传统育种利用自然等位基因适应特征的遗传多样性来改良植物抗旱性,基因组精准编辑工具和全基因组关联分析(Genome-wide Association Study,GWAS)等新技术的出现,在挖掘可提高抗旱性和产量的等位基因方面具有巨大应用潜力.生物刺激素中,一些小肽或激素激动剂的鉴定有助于精细调控干旱响应,在提高抗旱性同时保持了作物产量.这些技术的应用转化可为作物生产提供应对干旱的新策略(Gupta et al.,2020). ...
... 第二,根据气候资源特点,调整种植结构和种植制度以趋利避害.在半干旱地区通过实行“压夏扩秋”(即压缩春小麦、春玉米等春季作物种植比例)措施,扩大适应干旱能力较强且与降雨同季的马铃薯、大豆等秋季作物种植面积,应对干旱的季节变化特征.在以雨养农业为主的陇东黄土高原区,农业生产中稳定冬小麦种植面积,增加玉米面积,发展马铃薯、豆类、糜子、谷子等抗旱性较强的作物(邓振镛等,2007;张强等,2012b;肖国举和李裕,2012).合理安排和调整作物种植面积和布局,加强水热资源的合理开发利用和管理,变被动抗旱为主动抗旱,管好、用好当地水资源,充分利用大气降水(张强等,2012b;王润元等,2015;鲍文中和周广胜,2017). ...
... ).合理安排和调整作物种植面积和布局,加强水热资源的合理开发利用和管理,变被动抗旱为主动抗旱,管好、用好当地水资源,充分利用大气降水(张强等,2012b;王润元等,2015;鲍文中和周广胜,2017). ...
... 第三,在水资源缺乏的灌溉区和干旱山区推行水旱并举方略,逐步建立灌溉农业、旱地农业、设施农业和半旱地农业并存的农业用水新格局(山仑,2011),在黄淮海地区已有主动采用半旱地农业的实例,如山东恒台县采用的低定额灌溉条件下保持高产已初见成效,节水灌溉、少灌的半旱地轮作体系,也是比较成功的经验之一(山仑,2011).灌溉区调整灌溉模式,采用滴灌、喷灌等节水灌溉技术.在旱作农业区,根据农作物各生育期耗水亏损值进行分阶段、分次补充灌溉,在有条件的地区,基于物联网技术进行精细化灌溉和调控用水.开发农业用水新水源,如垄沟集雨、保墒集水、蓄集雨水、再生水、凝结水等都可作为补充水源供给农田,也可通过开发空中水资源补充陆地水资源不足(山仑,2011;张强等,2012b;肖国举和李裕,2012;王润元等,2015;熊友才和李凤民,2014). ...
... 第四,因地制宜推广保护性耕作措施,可同时起到保水、保土、培肥、增产的综合作用,如垄沟种植、垄沟径流集水、地膜覆盖、秸秆覆盖、免耕栽培等技术.采取多种形式的带状间作为中心的保护性耕作技术,缓解气候变暖加剧引起的水资源供求矛盾.推行麦类等条播作物与马铃薯、玉米、大豆等穴播作物或主粮作物与畜草合理间作、轮作等技术;合理套作,增加复种指数,提高耕地利用效率,如小麦套种马铃薯、小麦套种玉米等,可不同程度地提高作物产量,相当于在半干旱偏旱区的旱作农田上实现一年二熟(山仑,2011;张强等,2012a;张强等,2012b;肖国举和李裕,2012;鲍文中和周广胜,2017;Zhao et al.,2012;Zhao et al.,2014). ...
中国西北地区干旱气象灾害监测预警与减灾技术研究进展及其展望
2
2015
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
中国干旱事件成因和变化规律的研究进展与展望
1
2020
... 农业干旱是农作物生长发育过程中,因降雨不足或长期无雨造成大气干旱、土壤缺水,致使作物得不到适时适量的水分补给,无法满足正常需水,影响其光合过程及生物量积累,最终造成农作物减产甚至绝收.农业干旱是影响农业生产的重要因素,也是农业生产面临的最大风险之一(王春乙,2007;肖国举和李裕,2012;张强等,2017;Mirzabaev et al.,2023).农业生产决定着粮食产量的稳定,是国家战略性、基础性核心产业,农作物种植是农业的“芯片”,是促进农业长期稳定发展,保障国家粮食安全的根本.干旱缺水直接影响农作物的生长发育和产量形成,造成粮食产量下降、品质降低,从而增加粮食生产供给的不稳定性和风险,直接威胁粮食安全,成为制约社会经济发展的重要因素之一(姚玉璧等,2013;张强等,2008;张强等,2020;IPCC,2007;IPCC,2022;Mirzabaev et al.,2023),长期以来农业旱灾对粮食安全的影响是学者和政府部门关注的重点和焦点.农业干旱对作物的影响从微观到宏观表现在细胞、生理和形态等不同层次水平上,其影响程度取决于干旱开始时间、发展和持续时间、严重程度及农作物种类和品种、所处生育期等因素(赵鸿等,2016;Ulrich et al.,2019;Gupta et al.,2020). ...
中国南方干旱灾害风险特征及其防控技术对策
4
2017
... 受全球气候变化和经济快速发展的影响,全球干旱呈现多发、加重趋势,中亚地区65%的区域表现为干旱程度加剧、强度持续增强,2022年全球极端干旱问题尤为突出(李稚等,2022;张强,2022;Metz et al.,2007;Li et al.,2015;IPCC,2022).随着全球气候变暖,水循环加快,陆地蒸散发和储水量减少,干旱半干旱区不断扩张,干旱持续加剧,干旱风险进一步加大,且存在较大的区域差异(张强等,2017;Salinger et al.,2005;Cook et al.,2007;Metz et al.,2007;Huang et al.,2016).联合国《2022年全球干旱数字》报告指出,自2000年以来全球干旱次数增加了29%,全球处于高温和干旱管理的“十字路口”(UNCCD, 2022).预计到2100年,极端干旱地区的干旱次数和强度将增加1%~30%(IPCC,2007). ...
... 农业干旱是农作物生长发育过程中,因降雨不足或长期无雨造成大气干旱、土壤缺水,致使作物得不到适时适量的水分补给,无法满足正常需水,影响其光合过程及生物量积累,最终造成农作物减产甚至绝收.农业干旱是影响农业生产的重要因素,也是农业生产面临的最大风险之一(王春乙,2007;肖国举和李裕,2012;张强等,2017;Mirzabaev et al.,2023).农业生产决定着粮食产量的稳定,是国家战略性、基础性核心产业,农作物种植是农业的“芯片”,是促进农业长期稳定发展,保障国家粮食安全的根本.干旱缺水直接影响农作物的生长发育和产量形成,造成粮食产量下降、品质降低,从而增加粮食生产供给的不稳定性和风险,直接威胁粮食安全,成为制约社会经济发展的重要因素之一(姚玉璧等,2013;张强等,2008;张强等,2020;IPCC,2007;IPCC,2022;Mirzabaev et al.,2023),长期以来农业旱灾对粮食安全的影响是学者和政府部门关注的重点和焦点.农业干旱对作物的影响从微观到宏观表现在细胞、生理和形态等不同层次水平上,其影响程度取决于干旱开始时间、发展和持续时间、严重程度及农作物种类和品种、所处生育期等因素(赵鸿等,2016;Ulrich et al.,2019;Gupta et al.,2020). ...
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 2012年夏季美国遭遇了56 a来最严重旱灾,导致严重的粮食产量下降、品质降低和价格猛涨,其中玉米和小麦价格分别上涨60%和26%,引发了全球性的粮食危机.同年,受干旱气候影响,俄罗斯、乌克兰、中国等国家小麦、玉米主产区减产,也引起了不同程度的粮食减产和价格上涨(张强等,2014).2010年夏季俄罗斯遭受了130 a来最严重的干旱,导致近1/3的农作物受灾,因粮食减产而暂停小麦、玉米等农产品出口(https://www.docin.com/p-1435663470.html). 2016年越南南部遭遇近百年来的严重干旱,干旱对水稻影响尤为严重,湄公河三角洲区域农业受灾严重(张强等,2017).预计到21世纪末,气候变化将对非洲和中南美洲谷物产生负面影响,叠加物候影响,这将势必改变当前全球作物适宜种植区(https://m.thepaper.cn/baijiahao_20016300).2022年11月15日,联合国秘书长在二十国集团峰会上表示,如果不采取协调行动,今年的“粮食负担能力危机”可能在2023年变成严重的全球粮食短缺问题,并呼吁现在就共同行动起来,防止明年发生“严重的粮食灾难”(https://news.un.org/zh/story/2022/11/1112462). ...
农业干旱预警研究现状及发展趋势
2
2011
... 2006年,美国开始建设协调综合的国家干旱早期预警系统(Drought Early Warning System, DEWS),随后又建成了国家综合干旱信息系统(National Integrated Drought Information System,NIDIS);2007年,地球观测部计划未来建立全球干旱早期预警系统(Global Drought Early Warning System,GDEWS).美国利用MODIS(Moderate Resolution Imaging Spectroradiometer)产品监测全球干旱及旱灾造成的作物减产分布,用户可通过网络界面查询预先定置区域的数据并分析农作物长势和旱情(范锦龙等,2014).此外,美国国家干旱减灾中心(National Drought Mitigation Center,NDMC)联合美国农业部(The United States Department of Agriculture,USDA)等相关部门研发包括气象指标、土壤墒情指标的干旱监测系统(张晓煜等,2011;纪瑞鹏等,2019).中国风云卫星也具有监测全球干旱的能力,农业监测是其一个关键的应用领域,如FY-3A和FY-3B携带的可见光近红外扫描辐射计(Visible Infrared Radiometer,VIRR)和中分辨率成像仪(Medium Resolution Spectral Imager,MERSI)是农业监测的关键传感器,可监测不同尺度干旱灾害的发生发展(张晓煜等,2011;范锦龙等,2014).此外,中国国家气候中心也开展了干旱监测、预测预警和影响评估业务,发布全球旱涝指数、中国气象干旱综合指数、土壤相对湿度等系列干旱产品和公告,为抗旱减灾提供了重要的决策服务信息(范锦龙等,2014).在国家科技部支持下,气象、农业等部门联合运用气象灾害预测预报技术进行重大农业气象灾害监测预测和预警技术的攻关研发(陈德亮,2012;纪瑞鹏等,2019). ...
... ).中国风云卫星也具有监测全球干旱的能力,农业监测是其一个关键的应用领域,如FY-3A和FY-3B携带的可见光近红外扫描辐射计(Visible Infrared Radiometer,VIRR)和中分辨率成像仪(Medium Resolution Spectral Imager,MERSI)是农业监测的关键传感器,可监测不同尺度干旱灾害的发生发展(张晓煜等,2011;范锦龙等,2014).此外,中国国家气候中心也开展了干旱监测、预测预警和影响评估业务,发布全球旱涝指数、中国气象干旱综合指数、土壤相对湿度等系列干旱产品和公告,为抗旱减灾提供了重要的决策服务信息(范锦龙等,2014).在国家科技部支持下,气象、农业等部门联合运用气象灾害预测预报技术进行重大农业气象灾害监测预测和预警技术的攻关研发(陈德亮,2012;纪瑞鹏等,2019). ...
土壤水分胁迫对玉米生长发育的影响研究进展
2
2012
... 植物生长调节剂(外用)和植物激素(内用)是影响植物生理过程的物质,其浓度很低时起调节作用,如脱落酸、乙烯、生长素、赤霉素、细胞分裂素等(Morgan,1991;康书江等,1997).在分子和生化水平上,植物通过在转录和蛋白表达过程中改变调节反应来感知和响应干旱胁迫,从而影响生化途径和代谢过程,进而影响生理和发育过程(Mane et al.,2008;Vasquez-Robinet et al.,2008;Zinselmeier et al.,1995).在土壤干旱条件下植物通常会产生脱落酸(Abscisic Acid,ABA),它是一种主要的化学根-芽胁迫信号(Davies and Zhang,1991),是通过激活对干旱响应的基因表达而表征细胞反应的一个关键信号(Chaves et al.,2003).ABA含量可作为植物抗旱性的评价指标之一,其与籽粒的生长速率有关.当土壤干旱时失水的根系产生根源信号ABA,通过木质部输送到地上部调节气孔开闭(Davies and Zhang,1991),作物根冠、叶片、花、籽粒生长进程中ABA含量明显上升(赵鸿等,2008;张玉书等,2012;Obidiegwu et al,2015),且随着干旱胁迫程度的加剧而增加,如受到水分胁迫时小麦籽粒生长减慢、结实率降低,玉米胚乳细胞分化率降低.乙烯(Ethylene)也是干旱胁迫下植物体产生的激素,在干旱感知时乙烯通过蒸腾流将其他激素信号与ABA一起发送到嫩芽中.干旱引发的乙烯释放过量,会引起小麦、玉米等籽粒产量下降(赵鸿等,2008;Obidiegwu et al.,2015). ...
... 叶片对CO2的同化主要受气孔关闭、膜损伤和各种酶活性的干扰,尤其是CO2固定酶和三磷酸腺苷(ATP)合成酶,通过光呼吸途径增强的代谢物通量增加组织的氧化负荷,该过程产生活性氧,干旱胁迫下活性氧对生物大分子的损伤是影响植物生长的主要因素之一(Tezara et al.,1999;Farooq et al.,2009).干旱缺水时,作物体内活性氧的积累受到破坏,抗氧化酶活性降低,对活性氧的清除能力下降,致使作物避免或减轻受氧化伤害(赵鸿等,2008;Obidiegwu et al.,2015).轻度干旱可引起叶片中过氧化氢酶(Catalase,CAT)活性升高,而重度干旱下CAT活性降低.随着干旱胁迫的持续,叶片中过氧化物酶(Peroxidase,POD)活性渐增,丙二醛(Malonic Dialdehyde,MDA)含量增大,膜脂、蛋白质和核酸氧化加剧,明显抑制了植株的生长发育(张玉书等,2012;Obidiegwu et al.,2015).马铃薯在遭遇不同程度干旱胁迫时,随着胁迫程度的增加,植株体内超氧化物歧化酶(Superoxide Dismutase,SOD)活性下降,马铃薯受自由基损伤加重,无法起到保护细胞、去除活性氧自由基的作用,细胞受伤害增大、植株老化加速.在极度干旱胁迫下,当活性氧浓度超出清除剂的潜力时,干旱对不同植物细胞包括脂质、蛋白质和脱氧核糖核酸(Deoxyribo Nucleic Acid,DNA)造成不可修复的损害,导致植物细胞死亡(Obidiegwu et al.,2015;Gupta et al.,2020). ...
土壤干旱对作物生长过程和产量影响的研究进展
4
2008
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
... 植物生长调节剂(外用)和植物激素(内用)是影响植物生理过程的物质,其浓度很低时起调节作用,如脱落酸、乙烯、生长素、赤霉素、细胞分裂素等(Morgan,1991;康书江等,1997).在分子和生化水平上,植物通过在转录和蛋白表达过程中改变调节反应来感知和响应干旱胁迫,从而影响生化途径和代谢过程,进而影响生理和发育过程(Mane et al.,2008;Vasquez-Robinet et al.,2008;Zinselmeier et al.,1995).在土壤干旱条件下植物通常会产生脱落酸(Abscisic Acid,ABA),它是一种主要的化学根-芽胁迫信号(Davies and Zhang,1991),是通过激活对干旱响应的基因表达而表征细胞反应的一个关键信号(Chaves et al.,2003).ABA含量可作为植物抗旱性的评价指标之一,其与籽粒的生长速率有关.当土壤干旱时失水的根系产生根源信号ABA,通过木质部输送到地上部调节气孔开闭(Davies and Zhang,1991),作物根冠、叶片、花、籽粒生长进程中ABA含量明显上升(赵鸿等,2008;张玉书等,2012;Obidiegwu et al,2015),且随着干旱胁迫程度的加剧而增加,如受到水分胁迫时小麦籽粒生长减慢、结实率降低,玉米胚乳细胞分化率降低.乙烯(Ethylene)也是干旱胁迫下植物体产生的激素,在干旱感知时乙烯通过蒸腾流将其他激素信号与ABA一起发送到嫩芽中.干旱引发的乙烯释放过量,会引起小麦、玉米等籽粒产量下降(赵鸿等,2008;Obidiegwu et al.,2015). ...
... ),且随着干旱胁迫程度的加剧而增加,如受到水分胁迫时小麦籽粒生长减慢、结实率降低,玉米胚乳细胞分化率降低.乙烯(Ethylene)也是干旱胁迫下植物体产生的激素,在干旱感知时乙烯通过蒸腾流将其他激素信号与ABA一起发送到嫩芽中.干旱引发的乙烯释放过量,会引起小麦、玉米等籽粒产量下降(赵鸿等,2008;Obidiegwu et al.,2015). ...
... 叶片对CO2的同化主要受气孔关闭、膜损伤和各种酶活性的干扰,尤其是CO2固定酶和三磷酸腺苷(ATP)合成酶,通过光呼吸途径增强的代谢物通量增加组织的氧化负荷,该过程产生活性氧,干旱胁迫下活性氧对生物大分子的损伤是影响植物生长的主要因素之一(Tezara et al.,1999;Farooq et al.,2009).干旱缺水时,作物体内活性氧的积累受到破坏,抗氧化酶活性降低,对活性氧的清除能力下降,致使作物避免或减轻受氧化伤害(赵鸿等,2008;Obidiegwu et al.,2015).轻度干旱可引起叶片中过氧化氢酶(Catalase,CAT)活性升高,而重度干旱下CAT活性降低.随着干旱胁迫的持续,叶片中过氧化物酶(Peroxidase,POD)活性渐增,丙二醛(Malonic Dialdehyde,MDA)含量增大,膜脂、蛋白质和核酸氧化加剧,明显抑制了植株的生长发育(张玉书等,2012;Obidiegwu et al.,2015).马铃薯在遭遇不同程度干旱胁迫时,随着胁迫程度的增加,植株体内超氧化物歧化酶(Superoxide Dismutase,SOD)活性下降,马铃薯受自由基损伤加重,无法起到保护细胞、去除活性氧自由基的作用,细胞受伤害增大、植株老化加速.在极度干旱胁迫下,当活性氧浓度超出清除剂的潜力时,干旱对不同植物细胞包括脂质、蛋白质和脱氧核糖核酸(Deoxyribo Nucleic Acid,DNA)造成不可修复的损害,导致植物细胞死亡(Obidiegwu et al.,2015;Gupta et al.,2020). ...
马铃薯对土壤水分胁迫响应的研究进展
2
2018
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 在农业干旱监测中,农业干旱指标主要有气象指标、土壤墒情指标、作物生理生态指标和一些其他指标.气象指标包括降水距平百分数、连续无有效降水日数、湿度指标和Z指数等(孙荣强,1994;姚玉璧等,2013;国家气象中心等,2015;Zhang et al.,2009).土壤墒情指标包括土壤相对湿度、土壤有效水分存储量、土壤水分旱灾指数等(国家气象中心等,2015;Hollinger et al.,1993;Dai et al.,2004).作物生理生态指标主要是一些基于植株生长和光合特性的生理参数,用来衡量植株水分亏缺状况,如光合速率、蒸腾速率、气孔导度、复水后的光合恢复、叶水势、茎秆水势、叶绿素、叶片相对含水量、叶片扩展速率、作物水分胁迫指数、冠层温度、块茎碳同位素鉴别等(姚玉璧等,2013;张强等,2014;赵鸿等,2018;纪瑞鹏等,2019;Jefferies,1993;Medrano et al.,2002;Zakaluk and Ranjan,2006;Ricardo,2012;Ramirez et al.,2016),这些基于植株的监测指标更加方便、适时,但有些地区由于没有专用测量仪器而导致部分指标不能广泛应用. ...
粮食作物对高温干旱胁迫的响应及其阈值研究进展与展望
6
2016
... 农业干旱是农作物生长发育过程中,因降雨不足或长期无雨造成大气干旱、土壤缺水,致使作物得不到适时适量的水分补给,无法满足正常需水,影响其光合过程及生物量积累,最终造成农作物减产甚至绝收.农业干旱是影响农业生产的重要因素,也是农业生产面临的最大风险之一(王春乙,2007;肖国举和李裕,2012;张强等,2017;Mirzabaev et al.,2023).农业生产决定着粮食产量的稳定,是国家战略性、基础性核心产业,农作物种植是农业的“芯片”,是促进农业长期稳定发展,保障国家粮食安全的根本.干旱缺水直接影响农作物的生长发育和产量形成,造成粮食产量下降、品质降低,从而增加粮食生产供给的不稳定性和风险,直接威胁粮食安全,成为制约社会经济发展的重要因素之一(姚玉璧等,2013;张强等,2008;张强等,2020;IPCC,2007;IPCC,2022;Mirzabaev et al.,2023),长期以来农业旱灾对粮食安全的影响是学者和政府部门关注的重点和焦点.农业干旱对作物的影响从微观到宏观表现在细胞、生理和形态等不同层次水平上,其影响程度取决于干旱开始时间、发展和持续时间、严重程度及农作物种类和品种、所处生育期等因素(赵鸿等,2016;Ulrich et al.,2019;Gupta et al.,2020). ...
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... ;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
适应气候变化的国际行动和农业措施研究进展
5
2016
... 我国是一个农业大国,也是旱灾频发的国家,旱灾影响范围广、历时长、危害重.据统计,我国每年因干旱受灾的农田面积多达2 600万hm2,粮食减产高达50%~75%(姚玉璧等,2013;张强等,2014).近年来,受气候变化和水资源短缺的影响,我国农作物受旱、成灾和绝收面积不断扩大,在北方干旱形势依然严峻的情况下南方干旱出现明显增加和加重趋势,新旱区逐渐扩展,局部性、区域性干旱灾害频频出现,损失巨大(张强等,2014;康蕾和张红旗,2014;倪深海等,2022;张强,2022),严重威胁着国家的粮食安全和生态安全(王春乙,2007;张强等,2012a;张强等,2012b;肖国举和李裕,2012;康蕾和张红旗,2014;周广胜等,2016;Fu et al.,2013),已成为制约社会经济可持续发展的最主要因素,给我国农业生产和抗旱减灾工作带来前所未有的挑战和困难(吕娟等,2011;张强等,2014;Huang et al.,2016). ...
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 第六,加强农业干旱监测预测预警,强化防旱减灾应对准备.在出现旱情的重点地区加强土壤墒情监测和定点调查,构建区域信息共享平台,及时掌握旱情发展蔓延动态;加快构建农业农村气象灾害预警信息发布系统,搭建多种渠道的气象服务信息传播平台,建立气象灾害风险评估体系,研发基于影响的农业干旱监测预测预警技术,针对农业生产中重要环节和农作物生长发育的水分需求,提升农业防旱减灾服务能力,定期发布土壤墒情监测公报、干旱监测公报、农业气象旬(周)报等农业气象服务产品(姚国章和袁敏,2010;张强等,2014;周广胜等,2016).总之,在确保国家和区域(或省、市、区)粮、棉、油需求安全前提下,充分利用水热资源优势,压缩高耗水作物和品种的种植面积,实行农业补贴政策,实现农业经济和水资源安全协调发展(鲍文中和周广胜,2017). ...
... 农业防旱减灾是一项复杂的社会系统工程,涉及旱灾发生前的监测、预报、预警和防御,以及灾害发生时抗灾、灾后救灾和重建等多个方面.依照“工程措施与非工程措施相结合”的原则,在加强水利工程设施建设的同时,还需强化政策法规、监测预警、旱中保障等抗旱非工程措施,构建旱情监测预警和决策系统,提高抗旱减灾工作的科学化、现代化和规范化水平(姚国章和袁敏,2010;周广胜等,2016).鉴于旱灾对农业生产影响较重,急需建立一个科学有效的农业旱灾保障机制,但目前我国这种保障机制仍较滞后,相应的农业保险等项目发展缓慢(姚国章和袁敏,2010;周广胜等,2016).我国在干旱灾害管理方面的法律法规和政策尚有待健全,需借鉴欧美等发达国家经验,由被动的应急抗旱向主动的防旱模式转变(毛晓华,2020;屈艳萍等,2013).同时,我国在各类干旱(包括气象、农业、水文等)监测方面已开展了大量研究工作,但缺乏各部门间的有效协作机制,即气象、水文、农情、工业取水用水和相应供水部门之间的信息共享和联防联动.干旱整体评价指标构建上还需借鉴国外的干旱综合评价指标体系(康天军和李军波,2018;陈滢,2020).未来,我国还需制定更为详细的干旱灾害防御规划,让公众参与到干旱防灾、减灾工作中,构建全民参与的全社会防灾减灾新格局. ...
... ;周广胜等,2016).我国在干旱灾害管理方面的法律法规和政策尚有待健全,需借鉴欧美等发达国家经验,由被动的应急抗旱向主动的防旱模式转变(毛晓华,2020;屈艳萍等,2013).同时,我国在各类干旱(包括气象、农业、水文等)监测方面已开展了大量研究工作,但缺乏各部门间的有效协作机制,即气象、水文、农情、工业取水用水和相应供水部门之间的信息共享和联防联动.干旱整体评价指标构建上还需借鉴国外的干旱综合评价指标体系(康天军和李军波,2018;陈滢,2020).未来,我国还需制定更为详细的干旱灾害防御规划,让公众参与到干旱防灾、减灾工作中,构建全民参与的全社会防灾减灾新格局. ...
美国的干旱危机处理
1
2009
... 国际干旱减灾中心(International Centre for Drought Mitigation,IDMC)提出应对农业干旱的建议:首先要开展农业(田)灌溉效率的研究,鼓励安装节水灌溉设施,提供最优的农业水管理及灌溉设施维护措施;其次,鼓励各种节水农业灌溉新技术,并在干旱期为农业种植方案提供指导;第三,做好农业干旱监测,提供实时的土壤湿度和土壤水分蒸散发信息以及灌溉作业和安排;最后,面向农民推动农业/作物保险项目的发展(朱增勇和聂凤英,2009). ...
Growth and yield components of wheat under water stress of different growth stages
4
2011
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
Coordinated regulation of photosynthesis in rice increases yield and tolerance to environmental stress
1
2014
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
Meteorological drought-policy statement
1
1997
... 干旱灾害(简称“旱灾”),是指一段时间内降水量比常年明显偏少,造成某一地区经济社会活动(尤其是农业生产)和人类生活遭受较大危害的现象,普遍存在于世界各地,频繁发生于不同时期(Wilhite,2000;Li et al.,2015).旱灾是气象干旱、农业干旱、水文干旱、社会经济干旱和生态干旱之间的一种链状传递和转化过程,一个完整的干旱事件包括干旱历时、严重程度和平均强度(姚玉璧等,2013;张强等,2014;Heffernan,2013).全球每年因干旱导致的经济损失达60~80亿美元,2021年甚至高达121亿美元(American Meteorological Society, 1997;应急管理部-教育部减灾与应急管理研究院等,2022),远高于其他气象灾害造成的损失(张强等,2014;Wilhite,2000),受到高度关注(王春乙,2007;吕娟等,2011;Turner et al., 2011;Gupta et al.,2020). ...
Evaluation of drought indices based on thermal remote sensing of evapotranspiration over the Continental United States
1
2011
... 卫星遥感、地面遥感等探测技术的发展,为农作物旱情监测开辟了一条新途径.目前,国内外学者相继研发了多个农业干旱遥感监测指数,主要有以下四大类:(1)土壤水分指数,如表观热惯量(Apparent Thermal Inertia,ATI)、微波反演土壤水分等;(2)作物形态及绿度指数,如归一化植被指数(Normalized Difference Vegetation Index,NDVI)、植被状态指数(Vegetation Condition Index,VCI)、标准植被指数(Standardized Vegetation Index,SVI)等;(3)冠层温度指数,包括归一化温度指数(Normalized Difference Temperature Index,NDTI)、作物水分胁迫指数(Crop Water Stress Index,CWSI)、温度植被干旱指数(Temperature Vegetation Dryness Index,TVDI)、植被供水指数(Vegetation Supply Water Index,VSWI)等;(4)植被水分指数,包括全球植被水分指数(Global Vegetation Moisture Index,GVMI)、归一化差异水分指数(Normalized Difference Water Index,NDWI)等.这些指数各有优缺点,基于作物光谱信息,结合地面站点观测数据,可对大范围农作物干旱进行监测和评估,取得了较好的效果(李柏贞和周广胜,2014;纪瑞鹏等,2019;Anderson,2011). ...
Inhibition of photosynthesis in olive trees (Olea europaea L.) during water stress and rewatering
1
1996
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
Plant breeding and drought in C3 cereals: what should we breed for?
2
2002
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
Chlorophyll fluorescence: a probe of photosynthesis in vivo
1
2008
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
1
2012
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
Proline-rich cell wall proteins accumulate in growing regions and phloem tissue in response to water deficit in common bean seedlings
3
2007
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
Divergent strategies displayed by potato (Solanum tuberosum L.) cultivars to cope with soil drought
2
2018
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
Photosynthesis at low water potentials
1
1976
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
Grain yields with limited water
3
2004
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
Understanding plant responses to drought-from genes to the whole plant
3
2003
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
... 植物生长调节剂(外用)和植物激素(内用)是影响植物生理过程的物质,其浓度很低时起调节作用,如脱落酸、乙烯、生长素、赤霉素、细胞分裂素等(Morgan,1991;康书江等,1997).在分子和生化水平上,植物通过在转录和蛋白表达过程中改变调节反应来感知和响应干旱胁迫,从而影响生化途径和代谢过程,进而影响生理和发育过程(Mane et al.,2008;Vasquez-Robinet et al.,2008;Zinselmeier et al.,1995).在土壤干旱条件下植物通常会产生脱落酸(Abscisic Acid,ABA),它是一种主要的化学根-芽胁迫信号(Davies and Zhang,1991),是通过激活对干旱响应的基因表达而表征细胞反应的一个关键信号(Chaves et al.,2003).ABA含量可作为植物抗旱性的评价指标之一,其与籽粒的生长速率有关.当土壤干旱时失水的根系产生根源信号ABA,通过木质部输送到地上部调节气孔开闭(Davies and Zhang,1991),作物根冠、叶片、花、籽粒生长进程中ABA含量明显上升(赵鸿等,2008;张玉书等,2012;Obidiegwu et al,2015),且随着干旱胁迫程度的加剧而增加,如受到水分胁迫时小麦籽粒生长减慢、结实率降低,玉米胚乳细胞分化率降低.乙烯(Ethylene)也是干旱胁迫下植物体产生的激素,在干旱感知时乙烯通过蒸腾流将其他激素信号与ABA一起发送到嫩芽中.干旱引发的乙烯释放过量,会引起小麦、玉米等籽粒产量下降(赵鸿等,2008;Obidiegwu et al.,2015). ...
Stakeholder coinquiries on drought impacts, monitoring and early warning systems
1
2016
... 美国、澳大利亚等受干旱影响较重的国家,在过去30 a间,率先摒弃了被动应急抗旱的方式,积极采取主动的干旱风险管理模式,即预警防旱早于应急救援(Collins et al.,2016),有效应对可能发生的干旱灾害(Fu et al.,2013;Hervás-Gámez and Delgado-Ramos,2020).具体包括3个方面:首先,通过制定抗旱预案制度、发布国家和区域干旱政策、提高干旱监测预警能力、加强干旱灾害风险评估等手段,增强全社会防御干旱的能力(Wilhite et al.,2007;Peters-lidard et al.,2021;Steinemann et al.,2015;Zarei et al.,2020);其次,在干旱易发区域或干旱发生后,采取强化各部门需水管理、普及淡水资源化以及提高水分利用效率措施,如种植抗旱作物、农业节水灌溉、废水再利用及微咸水灌溉等,也可采用工程抗旱、生物抗旱等措施尽可能降低旱灾造成的损失(WMO and GWP,2014);第三,对于干旱造成损失的,政府制定相应政策评估受损,给予救济金补贴,让受灾严重的群体尽快恢复生产生活(WMO and GWP,2017).此外,澳大利亚还利用地下含水层储水-回灌技术发展农业节水灌溉,即把冬天地表多余的水资源通过压力储存到地下100 m左右的含水层,遇到干旱月份再抽出来进行农田灌溉;通过地理信息系统(Geographic Information System,GIS)、遥感系统(Remote Sensing,RS)、全球定位系统(Global Position System,GPS)等3S系统和制图(Mapping)、监控(Monitoring)、管理(Management)系统等3M系统进行节水灌溉,以实现节约水资源的目的(https://world.qianlong.com/2022/0830/7575142.shtml). ...
North American drought: reconstructions, causes, and consequences
1
2007
... 受全球气候变化和经济快速发展的影响,全球干旱呈现多发、加重趋势,中亚地区65%的区域表现为干旱程度加剧、强度持续增强,2022年全球极端干旱问题尤为突出(李稚等,2022;张强,2022;Metz et al.,2007;Li et al.,2015;IPCC,2022).随着全球气候变暖,水循环加快,陆地蒸散发和储水量减少,干旱半干旱区不断扩张,干旱持续加剧,干旱风险进一步加大,且存在较大的区域差异(张强等,2017;Salinger et al.,2005;Cook et al.,2007;Metz et al.,2007;Huang et al.,2016).联合国《2022年全球干旱数字》报告指出,自2000年以来全球干旱次数增加了29%,全球处于高温和干旱管理的“十字路口”(UNCCD, 2022).预计到2100年,极端干旱地区的干旱次数和强度将增加1%~30%(IPCC,2007). ...
Stomatal and non-stomatal contribution in the decline in leaf net CO2 uptake during rapid water stress
1
1983
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
Yield, water use efficiency and nitrogen uptake in potato: influence of drought stress
1
1997
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
A global dataset of Palmer drought severity index for 1870-2002: relationship with soil moisture and effects of surface warming
2
2004
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 在农业干旱监测中,农业干旱指标主要有气象指标、土壤墒情指标、作物生理生态指标和一些其他指标.气象指标包括降水距平百分数、连续无有效降水日数、湿度指标和Z指数等(孙荣强,1994;姚玉璧等,2013;国家气象中心等,2015;Zhang et al.,2009).土壤墒情指标包括土壤相对湿度、土壤有效水分存储量、土壤水分旱灾指数等(国家气象中心等,2015;Hollinger et al.,1993;Dai et al.,2004).作物生理生态指标主要是一些基于植株生长和光合特性的生理参数,用来衡量植株水分亏缺状况,如光合速率、蒸腾速率、气孔导度、复水后的光合恢复、叶水势、茎秆水势、叶绿素、叶片相对含水量、叶片扩展速率、作物水分胁迫指数、冠层温度、块茎碳同位素鉴别等(姚玉璧等,2013;张强等,2014;赵鸿等,2018;纪瑞鹏等,2019;Jefferies,1993;Medrano et al.,2002;Zakaluk and Ranjan,2006;Ricardo,2012;Ramirez et al.,2016),这些基于植株的监测指标更加方便、适时,但有些地区由于没有专用测量仪器而导致部分指标不能广泛应用. ...
Root signals and the regulation of growth and development of plants in drying soil
2
1991
... 植物生长调节剂(外用)和植物激素(内用)是影响植物生理过程的物质,其浓度很低时起调节作用,如脱落酸、乙烯、生长素、赤霉素、细胞分裂素等(Morgan,1991;康书江等,1997).在分子和生化水平上,植物通过在转录和蛋白表达过程中改变调节反应来感知和响应干旱胁迫,从而影响生化途径和代谢过程,进而影响生理和发育过程(Mane et al.,2008;Vasquez-Robinet et al.,2008;Zinselmeier et al.,1995).在土壤干旱条件下植物通常会产生脱落酸(Abscisic Acid,ABA),它是一种主要的化学根-芽胁迫信号(Davies and Zhang,1991),是通过激活对干旱响应的基因表达而表征细胞反应的一个关键信号(Chaves et al.,2003).ABA含量可作为植物抗旱性的评价指标之一,其与籽粒的生长速率有关.当土壤干旱时失水的根系产生根源信号ABA,通过木质部输送到地上部调节气孔开闭(Davies and Zhang,1991),作物根冠、叶片、花、籽粒生长进程中ABA含量明显上升(赵鸿等,2008;张玉书等,2012;Obidiegwu et al,2015),且随着干旱胁迫程度的加剧而增加,如受到水分胁迫时小麦籽粒生长减慢、结实率降低,玉米胚乳细胞分化率降低.乙烯(Ethylene)也是干旱胁迫下植物体产生的激素,在干旱感知时乙烯通过蒸腾流将其他激素信号与ABA一起发送到嫩芽中.干旱引发的乙烯释放过量,会引起小麦、玉米等籽粒产量下降(赵鸿等,2008;Obidiegwu et al.,2015). ...
... ).ABA含量可作为植物抗旱性的评价指标之一,其与籽粒的生长速率有关.当土壤干旱时失水的根系产生根源信号ABA,通过木质部输送到地上部调节气孔开闭(Davies and Zhang,1991),作物根冠、叶片、花、籽粒生长进程中ABA含量明显上升(赵鸿等,2008;张玉书等,2012;Obidiegwu et al,2015),且随着干旱胁迫程度的加剧而增加,如受到水分胁迫时小麦籽粒生长减慢、结实率降低,玉米胚乳细胞分化率降低.乙烯(Ethylene)也是干旱胁迫下植物体产生的激素,在干旱感知时乙烯通过蒸腾流将其他激素信号与ABA一起发送到嫩芽中.干旱引发的乙烯释放过量,会引起小麦、玉米等籽粒产量下降(赵鸿等,2008;Obidiegwu et al.,2015). ...
Physiological responses of spring durum wheat cultivars to early-season drought in a Mediterranean environment
1
1998
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
The state of food insecurity in the world: eradicating world hunger—taking stock ten years after the World Food Summit
1
2006
... 联合国《2022年干旱数字》报告指出,干旱对世界农业生产产生了深远、广泛且低估了的影响,尤其对粮食安全的影响(Gupta et al.,2020;Mirzabaev et al.,2023).干旱引起的水分短缺会对农作物造成一定危害,导致作物产量降低、品质下降,甚至绝收.IPCC第六次评估报告(IPCC,2022)指出,21世纪气候变化对粮食产量的影响仍以负面为主,变幅随不同作物、时间、地区变化差异较大.气候变化使玉米、大豆、水稻、小麦产量每10 a分别减少2.3%、3.3%、0.7%、1.3%,据统计,每年因干旱造成的粮食减产超过其他因素造成的总和,在过去10 a中全球因干旱造成的作物产量损失约300亿美元(FAO,2018).图1展示了全球人口、可耕地面积、农业淡水需求量的过去、现在和未来(Gupta et al.,2020).可以看出,全球人口从1990年的50亿增加到目前的75亿多,预计到2050年将增加到97~100亿[图1(a)],同时全球粮食需求量也将加速增长,从如今的约21亿t增长到2050年的30亿t左右,届时预计有50亿人生活在缺水地区(FAO,2006;Gupta et al.,2020);尽管全球可耕地面积略有增加[图1(b)],考虑到目前每公顷作物产量的比率,预计需要额外增加100万hm2可耕土地才能确保粮食安全(Gupta et al.,2020);考虑到目前气候变化趋势,预计到2050年农业用水需求量可能增加1倍[图1(c)],淡水供应量将下降50%(Gleick,2000;Koncagül et al.,2020;Gupta et al.,2020). ...
The impact of disasters and crises on agriculture and food security
1
2018
... 联合国《2022年干旱数字》报告指出,干旱对世界农业生产产生了深远、广泛且低估了的影响,尤其对粮食安全的影响(Gupta et al.,2020;Mirzabaev et al.,2023).干旱引起的水分短缺会对农作物造成一定危害,导致作物产量降低、品质下降,甚至绝收.IPCC第六次评估报告(IPCC,2022)指出,21世纪气候变化对粮食产量的影响仍以负面为主,变幅随不同作物、时间、地区变化差异较大.气候变化使玉米、大豆、水稻、小麦产量每10 a分别减少2.3%、3.3%、0.7%、1.3%,据统计,每年因干旱造成的粮食减产超过其他因素造成的总和,在过去10 a中全球因干旱造成的作物产量损失约300亿美元(FAO,2018).图1展示了全球人口、可耕地面积、农业淡水需求量的过去、现在和未来(Gupta et al.,2020).可以看出,全球人口从1990年的50亿增加到目前的75亿多,预计到2050年将增加到97~100亿[图1(a)],同时全球粮食需求量也将加速增长,从如今的约21亿t增长到2050年的30亿t左右,届时预计有50亿人生活在缺水地区(FAO,2006;Gupta et al.,2020);尽管全球可耕地面积略有增加[图1(b)],考虑到目前每公顷作物产量的比率,预计需要额外增加100万hm2可耕土地才能确保粮食安全(Gupta et al.,2020);考虑到目前气候变化趋势,预计到2050年农业用水需求量可能增加1倍[图1(c)],淡水供应量将下降50%(Gleick,2000;Koncagül et al.,2020;Gupta et al.,2020). ...
Plant drought stress: effects, mechanisms and management
6
2009
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
... ;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
... ;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
... 叶片对CO2的同化主要受气孔关闭、膜损伤和各种酶活性的干扰,尤其是CO2固定酶和三磷酸腺苷(ATP)合成酶,通过光呼吸途径增强的代谢物通量增加组织的氧化负荷,该过程产生活性氧,干旱胁迫下活性氧对生物大分子的损伤是影响植物生长的主要因素之一(Tezara et al.,1999;Farooq et al.,2009).干旱缺水时,作物体内活性氧的积累受到破坏,抗氧化酶活性降低,对活性氧的清除能力下降,致使作物避免或减轻受氧化伤害(赵鸿等,2008;Obidiegwu et al.,2015).轻度干旱可引起叶片中过氧化氢酶(Catalase,CAT)活性升高,而重度干旱下CAT活性降低.随着干旱胁迫的持续,叶片中过氧化物酶(Peroxidase,POD)活性渐增,丙二醛(Malonic Dialdehyde,MDA)含量增大,膜脂、蛋白质和核酸氧化加剧,明显抑制了植株的生长发育(张玉书等,2012;Obidiegwu et al.,2015).马铃薯在遭遇不同程度干旱胁迫时,随着胁迫程度的增加,植株体内超氧化物歧化酶(Superoxide Dismutase,SOD)活性下降,马铃薯受自由基损伤加重,无法起到保护细胞、去除活性氧自由基的作用,细胞受伤害增大、植株老化加速.在极度干旱胁迫下,当活性氧浓度超出清除剂的潜力时,干旱对不同植物细胞包括脂质、蛋白质和脱氧核糖核酸(Deoxyribo Nucleic Acid,DNA)造成不可修复的损害,导致植物细胞死亡(Obidiegwu et al.,2015;Gupta et al.,2020). ...
... 渗透调节、渗透保护、抗氧化和清除防御系统是作物抗旱性的重要基础.作物表现出一系列抵御干旱胁迫的机制,主要包括通过增加扩散阻力来减少水分流失、通过深层根系增强水分吸收和有效利用、通过更小和多汁的叶片来减少蒸腾损失(Farooq et al.,2009;Supratim et al.,2016).在营养物质中,钾离子有助于渗透调节.低分子渗透液,包括甜菜碱、脯氨酸和其他氨基酸、有机酸和多元醇等,在干旱条件下能维持细胞功能.在分子水平上目前已经鉴定出若干个干旱响应基因和转录因子,如脱水响应元件结合基因、水通道蛋白、胚胎发育后期丰富的蛋白质和脱氢蛋白等(Farooq et al.,2009;Supratim et al.,2016). ...
... ).在营养物质中,钾离子有助于渗透调节.低分子渗透液,包括甜菜碱、脯氨酸和其他氨基酸、有机酸和多元醇等,在干旱条件下能维持细胞功能.在分子水平上目前已经鉴定出若干个干旱响应基因和转录因子,如脱水响应元件结合基因、水通道蛋白、胚胎发育后期丰富的蛋白质和脱氢蛋白等(Farooq et al.,2009;Supratim et al.,2016). ...
An overview of US state drought plans: crisis or risk management?
3
2013
... 我国是一个农业大国,也是旱灾频发的国家,旱灾影响范围广、历时长、危害重.据统计,我国每年因干旱受灾的农田面积多达2 600万hm2,粮食减产高达50%~75%(姚玉璧等,2013;张强等,2014).近年来,受气候变化和水资源短缺的影响,我国农作物受旱、成灾和绝收面积不断扩大,在北方干旱形势依然严峻的情况下南方干旱出现明显增加和加重趋势,新旱区逐渐扩展,局部性、区域性干旱灾害频频出现,损失巨大(张强等,2014;康蕾和张红旗,2014;倪深海等,2022;张强,2022),严重威胁着国家的粮食安全和生态安全(王春乙,2007;张强等,2012a;张强等,2012b;肖国举和李裕,2012;康蕾和张红旗,2014;周广胜等,2016;Fu et al.,2013),已成为制约社会经济可持续发展的最主要因素,给我国农业生产和抗旱减灾工作带来前所未有的挑战和困难(吕娟等,2011;张强等,2014;Huang et al.,2016). ...
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 美国、澳大利亚等受干旱影响较重的国家,在过去30 a间,率先摒弃了被动应急抗旱的方式,积极采取主动的干旱风险管理模式,即预警防旱早于应急救援(Collins et al.,2016),有效应对可能发生的干旱灾害(Fu et al.,2013;Hervás-Gámez and Delgado-Ramos,2020).具体包括3个方面:首先,通过制定抗旱预案制度、发布国家和区域干旱政策、提高干旱监测预警能力、加强干旱灾害风险评估等手段,增强全社会防御干旱的能力(Wilhite et al.,2007;Peters-lidard et al.,2021;Steinemann et al.,2015;Zarei et al.,2020);其次,在干旱易发区域或干旱发生后,采取强化各部门需水管理、普及淡水资源化以及提高水分利用效率措施,如种植抗旱作物、农业节水灌溉、废水再利用及微咸水灌溉等,也可采用工程抗旱、生物抗旱等措施尽可能降低旱灾造成的损失(WMO and GWP,2014);第三,对于干旱造成损失的,政府制定相应政策评估受损,给予救济金补贴,让受灾严重的群体尽快恢复生产生活(WMO and GWP,2017).此外,澳大利亚还利用地下含水层储水-回灌技术发展农业节水灌溉,即把冬天地表多余的水资源通过压力储存到地下100 m左右的含水层,遇到干旱月份再抽出来进行农田灌溉;通过地理信息系统(Geographic Information System,GIS)、遥感系统(Remote Sensing,RS)、全球定位系统(Global Position System,GPS)等3S系统和制图(Mapping)、监控(Monitoring)、管理(Management)系统等3M系统进行节水灌溉,以实现节约水资源的目的(https://world.qianlong.com/2022/0830/7575142.shtml). ...
The physiology of drought stress in grapevine: towards an integrative definition of drought tolerance
1
2020
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
DA SILVA J B, 1987. Effects of drought on primary photosynthetic processes of cotton leaves
1
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
1
2000
... 联合国《2022年干旱数字》报告指出,干旱对世界农业生产产生了深远、广泛且低估了的影响,尤其对粮食安全的影响(Gupta et al.,2020;Mirzabaev et al.,2023).干旱引起的水分短缺会对农作物造成一定危害,导致作物产量降低、品质下降,甚至绝收.IPCC第六次评估报告(IPCC,2022)指出,21世纪气候变化对粮食产量的影响仍以负面为主,变幅随不同作物、时间、地区变化差异较大.气候变化使玉米、大豆、水稻、小麦产量每10 a分别减少2.3%、3.3%、0.7%、1.3%,据统计,每年因干旱造成的粮食减产超过其他因素造成的总和,在过去10 a中全球因干旱造成的作物产量损失约300亿美元(FAO,2018).图1展示了全球人口、可耕地面积、农业淡水需求量的过去、现在和未来(Gupta et al.,2020).可以看出,全球人口从1990年的50亿增加到目前的75亿多,预计到2050年将增加到97~100亿[图1(a)],同时全球粮食需求量也将加速增长,从如今的约21亿t增长到2050年的30亿t左右,届时预计有50亿人生活在缺水地区(FAO,2006;Gupta et al.,2020);尽管全球可耕地面积略有增加[图1(b)],考虑到目前每公顷作物产量的比率,预计需要额外增加100万hm2可耕土地才能确保粮食安全(Gupta et al.,2020);考虑到目前气候变化趋势,预计到2050年农业用水需求量可能增加1倍[图1(c)],淡水供应量将下降50%(Gleick,2000;Koncagül et al.,2020;Gupta et al.,2020). ...
The physiology of plant responses to drought
17
2020
... 干旱灾害(简称“旱灾”),是指一段时间内降水量比常年明显偏少,造成某一地区经济社会活动(尤其是农业生产)和人类生活遭受较大危害的现象,普遍存在于世界各地,频繁发生于不同时期(Wilhite,2000;Li et al.,2015).旱灾是气象干旱、农业干旱、水文干旱、社会经济干旱和生态干旱之间的一种链状传递和转化过程,一个完整的干旱事件包括干旱历时、严重程度和平均强度(姚玉璧等,2013;张强等,2014;Heffernan,2013).全球每年因干旱导致的经济损失达60~80亿美元,2021年甚至高达121亿美元(American Meteorological Society, 1997;应急管理部-教育部减灾与应急管理研究院等,2022),远高于其他气象灾害造成的损失(张强等,2014;Wilhite,2000),受到高度关注(王春乙,2007;吕娟等,2011;Turner et al., 2011;Gupta et al.,2020). ...
... 农业干旱是农作物生长发育过程中,因降雨不足或长期无雨造成大气干旱、土壤缺水,致使作物得不到适时适量的水分补给,无法满足正常需水,影响其光合过程及生物量积累,最终造成农作物减产甚至绝收.农业干旱是影响农业生产的重要因素,也是农业生产面临的最大风险之一(王春乙,2007;肖国举和李裕,2012;张强等,2017;Mirzabaev et al.,2023).农业生产决定着粮食产量的稳定,是国家战略性、基础性核心产业,农作物种植是农业的“芯片”,是促进农业长期稳定发展,保障国家粮食安全的根本.干旱缺水直接影响农作物的生长发育和产量形成,造成粮食产量下降、品质降低,从而增加粮食生产供给的不稳定性和风险,直接威胁粮食安全,成为制约社会经济发展的重要因素之一(姚玉璧等,2013;张强等,2008;张强等,2020;IPCC,2007;IPCC,2022;Mirzabaev et al.,2023),长期以来农业旱灾对粮食安全的影响是学者和政府部门关注的重点和焦点.农业干旱对作物的影响从微观到宏观表现在细胞、生理和形态等不同层次水平上,其影响程度取决于干旱开始时间、发展和持续时间、严重程度及农作物种类和品种、所处生育期等因素(赵鸿等,2016;Ulrich et al.,2019;Gupta et al.,2020). ...
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... ;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 联合国《2022年干旱数字》报告指出,干旱对世界农业生产产生了深远、广泛且低估了的影响,尤其对粮食安全的影响(Gupta et al.,2020;Mirzabaev et al.,2023).干旱引起的水分短缺会对农作物造成一定危害,导致作物产量降低、品质下降,甚至绝收.IPCC第六次评估报告(IPCC,2022)指出,21世纪气候变化对粮食产量的影响仍以负面为主,变幅随不同作物、时间、地区变化差异较大.气候变化使玉米、大豆、水稻、小麦产量每10 a分别减少2.3%、3.3%、0.7%、1.3%,据统计,每年因干旱造成的粮食减产超过其他因素造成的总和,在过去10 a中全球因干旱造成的作物产量损失约300亿美元(FAO,2018).图1展示了全球人口、可耕地面积、农业淡水需求量的过去、现在和未来(Gupta et al.,2020).可以看出,全球人口从1990年的50亿增加到目前的75亿多,预计到2050年将增加到97~100亿[图1(a)],同时全球粮食需求量也将加速增长,从如今的约21亿t增长到2050年的30亿t左右,届时预计有50亿人生活在缺水地区(FAO,2006;Gupta et al.,2020);尽管全球可耕地面积略有增加[图1(b)],考虑到目前每公顷作物产量的比率,预计需要额外增加100万hm2可耕土地才能确保粮食安全(Gupta et al.,2020);考虑到目前气候变化趋势,预计到2050年农业用水需求量可能增加1倍[图1(c)],淡水供应量将下降50%(Gleick,2000;Koncagül et al.,2020;Gupta et al.,2020). ...
... 展示了全球人口、可耕地面积、农业淡水需求量的过去、现在和未来(Gupta et al.,2020).可以看出,全球人口从1990年的50亿增加到目前的75亿多,预计到2050年将增加到97~100亿[图1(a)],同时全球粮食需求量也将加速增长,从如今的约21亿t增长到2050年的30亿t左右,届时预计有50亿人生活在缺水地区(FAO,2006;Gupta et al.,2020);尽管全球可耕地面积略有增加[图1(b)],考虑到目前每公顷作物产量的比率,预计需要额外增加100万hm2可耕土地才能确保粮食安全(Gupta et al.,2020);考虑到目前气候变化趋势,预计到2050年农业用水需求量可能增加1倍[图1(c)],淡水供应量将下降50%(Gleick,2000;Koncagül et al.,2020;Gupta et al.,2020). ...
... ;Gupta et al.,2020);尽管全球可耕地面积略有增加[图1(b)],考虑到目前每公顷作物产量的比率,预计需要额外增加100万hm2可耕土地才能确保粮食安全(Gupta et al.,2020);考虑到目前气候变化趋势,预计到2050年农业用水需求量可能增加1倍[图1(c)],淡水供应量将下降50%(Gleick,2000;Koncagül et al.,2020;Gupta et al.,2020). ...
... 可耕土地才能确保粮食安全(Gupta et al.,2020);考虑到目前气候变化趋势,预计到2050年农业用水需求量可能增加1倍[图1(c)],淡水供应量将下降50%(Gleick,2000;Koncagül et al.,2020;Gupta et al.,2020). ...
... ;Gupta et al.,2020). ...
... 全球人口(a)、可耕地面积(b)和农业淡水需求量(c)的过去、现在和未来(Gupta et al.,
2020)
Past, present, and future of world population (a), arable land (b) and freshwater demand in agriculture (c)(Gupta et al., <xref ref-type="bibr" rid="b80">2020</xref>)Fig.1![]() 1.1.2 旱灾频发造成农作物产量下降、粮食价格上涨,粮食安全风险增大
1.1.2 旱灾频发造成农作物产量下降、粮食价格上涨,粮食安全风险增大2022年气候异常,北半球遭遇了大范围、史无前例的严重干旱,北美洲、欧洲、地中海地区、东北非地区以及中国南方地区都出现破纪录的极端酷热天气,许多地区经历了极端干旱(图2),对农业生产产生较大影响,大部分农作物有一定程度的减产.2022年9月联合国粮农组织发布的月报,将2022年全球谷物产量预测从7月初预测的27.92亿t下调至27.74亿t,比2021年预估产量低1.4%(https://xueqiu.com/4587623715/231014661).1964—2007年极端天气灾害造成全球粮食作物大面积减产,尤其是玉米、水稻、小麦,其中干旱和极端高温导致各国粮食减产9%~10%,发达国家的粮食产量损失比发展中国家高出8%~11%(纪瑞鹏等,2019;Lesk et al.,2016;Mirzabaev et al.,2023) ...
... Past, present, and future of world population (a), arable land (b) and freshwater demand in agriculture (c)(Gupta et al.,
2020)
Fig.1![]() 1.1.2 旱灾频发造成农作物产量下降、粮食价格上涨,粮食安全风险增大
1.1.2 旱灾频发造成农作物产量下降、粮食价格上涨,粮食安全风险增大2022年气候异常,北半球遭遇了大范围、史无前例的严重干旱,北美洲、欧洲、地中海地区、东北非地区以及中国南方地区都出现破纪录的极端酷热天气,许多地区经历了极端干旱(图2),对农业生产产生较大影响,大部分农作物有一定程度的减产.2022年9月联合国粮农组织发布的月报,将2022年全球谷物产量预测从7月初预测的27.92亿t下调至27.74亿t,比2021年预估产量低1.4%(https://xueqiu.com/4587623715/231014661).1964—2007年极端天气灾害造成全球粮食作物大面积减产,尤其是玉米、水稻、小麦,其中干旱和极端高温导致各国粮食减产9%~10%,发达国家的粮食产量损失比发展中国家高出8%~11%(纪瑞鹏等,2019;Lesk et al.,2016;Mirzabaev et al.,2023) ...
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
... 叶片对CO2的同化主要受气孔关闭、膜损伤和各种酶活性的干扰,尤其是CO2固定酶和三磷酸腺苷(ATP)合成酶,通过光呼吸途径增强的代谢物通量增加组织的氧化负荷,该过程产生活性氧,干旱胁迫下活性氧对生物大分子的损伤是影响植物生长的主要因素之一(Tezara et al.,1999;Farooq et al.,2009).干旱缺水时,作物体内活性氧的积累受到破坏,抗氧化酶活性降低,对活性氧的清除能力下降,致使作物避免或减轻受氧化伤害(赵鸿等,2008;Obidiegwu et al.,2015).轻度干旱可引起叶片中过氧化氢酶(Catalase,CAT)活性升高,而重度干旱下CAT活性降低.随着干旱胁迫的持续,叶片中过氧化物酶(Peroxidase,POD)活性渐增,丙二醛(Malonic Dialdehyde,MDA)含量增大,膜脂、蛋白质和核酸氧化加剧,明显抑制了植株的生长发育(张玉书等,2012;Obidiegwu et al.,2015).马铃薯在遭遇不同程度干旱胁迫时,随着胁迫程度的增加,植株体内超氧化物歧化酶(Superoxide Dismutase,SOD)活性下降,马铃薯受自由基损伤加重,无法起到保护细胞、去除活性氧自由基的作用,细胞受伤害增大、植株老化加速.在极度干旱胁迫下,当活性氧浓度超出清除剂的潜力时,干旱对不同植物细胞包括脂质、蛋白质和脱氧核糖核酸(Deoxyribo Nucleic Acid,DNA)造成不可修复的损害,导致植物细胞死亡(Obidiegwu et al.,2015;Gupta et al.,2020). ...
... 从分子生物学角度看,细胞水分损失是干旱胁迫的标志(Gupta et al.,2020).作物感知缺水信号并启动应对策略的能力被认为抗旱性.作物抗旱性是一个复杂性状,通常以避免或忍耐细胞脱水而防止水分流失来渡过干旱(Turner,1986;Machado and Paulsen,2001),主要表现为逃旱、避旱和耐旱等.逃旱是指作物通过调控生长发育期(提前或滞后水分敏感时段)来逃避干旱胁迫,一般发生在作物生长受阻之前;避旱,为减少水分损失,在干旱条件下保持高水势的能力,作物通过增加体内含水量避免组织损伤,涉及植株快速生长、叶片卷曲、叶面积减小、气孔开闭等(Morgan,1984;Turner,1986;Morgan,1991;Machado and Paulsen,2001;Gupta et al.,2020);耐旱是指组织内部含水量较低,干旱期间仍需保持生长耐力,在水分亏缺加重情况下通过保持充足的细胞膨胀来维持新陈代谢(Gupta et al.,2020). ...
... ;Gupta et al.,2020);耐旱是指组织内部含水量较低,干旱期间仍需保持生长耐力,在水分亏缺加重情况下通过保持充足的细胞膨胀来维持新陈代谢(Gupta et al.,2020). ...
... );耐旱是指组织内部含水量较低,干旱期间仍需保持生长耐力,在水分亏缺加重情况下通过保持充足的细胞膨胀来维持新陈代谢(Gupta et al.,2020). ...
... 第一,选择高产优质及抗旱、耐旱品种.培育与干旱相适应的作物新品种和优良品种,或者改良品种,特别是兼顾抗旱与水资源高效利用的品种(山仑,2011;张强等,2012b).开展水稻、玉米、小麦、马铃薯等作物新品种选育,对提高主粮生产能力具有重要作用.改良作物抗旱性的途径主要分为生物工程技术和生物激素鉴定两方面,在生物工程技术中传统育种利用自然等位基因适应特征的遗传多样性来改良植物抗旱性,基因组精准编辑工具和全基因组关联分析(Genome-wide Association Study,GWAS)等新技术的出现,在挖掘可提高抗旱性和产量的等位基因方面具有巨大应用潜力.生物刺激素中,一些小肽或激素激动剂的鉴定有助于精细调控干旱响应,在提高抗旱性同时保持了作物产量.这些技术的应用转化可为作物生产提供应对干旱的新策略(Gupta et al.,2020). ...
The dry facts
1
2013
... 干旱灾害(简称“旱灾”),是指一段时间内降水量比常年明显偏少,造成某一地区经济社会活动(尤其是农业生产)和人类生活遭受较大危害的现象,普遍存在于世界各地,频繁发生于不同时期(Wilhite,2000;Li et al.,2015).旱灾是气象干旱、农业干旱、水文干旱、社会经济干旱和生态干旱之间的一种链状传递和转化过程,一个完整的干旱事件包括干旱历时、严重程度和平均强度(姚玉璧等,2013;张强等,2014;Heffernan,2013).全球每年因干旱导致的经济损失达60~80亿美元,2021年甚至高达121亿美元(American Meteorological Society, 1997;应急管理部-教育部减灾与应急管理研究院等,2022),远高于其他气象灾害造成的损失(张强等,2014;Wilhite,2000),受到高度关注(王春乙,2007;吕娟等,2011;Turner et al., 2011;Gupta et al.,2020). ...
Are the modern drought management plans modern enough?The Guadalquivir River basin case in Spain
2
2020
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 美国、澳大利亚等受干旱影响较重的国家,在过去30 a间,率先摒弃了被动应急抗旱的方式,积极采取主动的干旱风险管理模式,即预警防旱早于应急救援(Collins et al.,2016),有效应对可能发生的干旱灾害(Fu et al.,2013;Hervás-Gámez and Delgado-Ramos,2020).具体包括3个方面:首先,通过制定抗旱预案制度、发布国家和区域干旱政策、提高干旱监测预警能力、加强干旱灾害风险评估等手段,增强全社会防御干旱的能力(Wilhite et al.,2007;Peters-lidard et al.,2021;Steinemann et al.,2015;Zarei et al.,2020);其次,在干旱易发区域或干旱发生后,采取强化各部门需水管理、普及淡水资源化以及提高水分利用效率措施,如种植抗旱作物、农业节水灌溉、废水再利用及微咸水灌溉等,也可采用工程抗旱、生物抗旱等措施尽可能降低旱灾造成的损失(WMO and GWP,2014);第三,对于干旱造成损失的,政府制定相应政策评估受损,给予救济金补贴,让受灾严重的群体尽快恢复生产生活(WMO and GWP,2017).此外,澳大利亚还利用地下含水层储水-回灌技术发展农业节水灌溉,即把冬天地表多余的水资源通过压力储存到地下100 m左右的含水层,遇到干旱月份再抽出来进行农田灌溉;通过地理信息系统(Geographic Information System,GIS)、遥感系统(Remote Sensing,RS)、全球定位系统(Global Position System,GPS)等3S系统和制图(Mapping)、监控(Monitoring)、管理(Management)系统等3M系统进行节水灌溉,以实现节约水资源的目的(https://world.qianlong.com/2022/0830/7575142.shtml). ...
Plant Responses to Abiotic Stress
1
2004
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
Combining ability of highland tropic adapted potato for tuber yield and yield components under drought
1
2017
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
A new soil moisture drought index for predicting crop yields
2
1993
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 在农业干旱监测中,农业干旱指标主要有气象指标、土壤墒情指标、作物生理生态指标和一些其他指标.气象指标包括降水距平百分数、连续无有效降水日数、湿度指标和Z指数等(孙荣强,1994;姚玉璧等,2013;国家气象中心等,2015;Zhang et al.,2009).土壤墒情指标包括土壤相对湿度、土壤有效水分存储量、土壤水分旱灾指数等(国家气象中心等,2015;Hollinger et al.,1993;Dai et al.,2004).作物生理生态指标主要是一些基于植株生长和光合特性的生理参数,用来衡量植株水分亏缺状况,如光合速率、蒸腾速率、气孔导度、复水后的光合恢复、叶水势、茎秆水势、叶绿素、叶片相对含水量、叶片扩展速率、作物水分胁迫指数、冠层温度、块茎碳同位素鉴别等(姚玉璧等,2013;张强等,2014;赵鸿等,2018;纪瑞鹏等,2019;Jefferies,1993;Medrano et al.,2002;Zakaluk and Ranjan,2006;Ricardo,2012;Ramirez et al.,2016),这些基于植株的监测指标更加方便、适时,但有些地区由于没有专用测量仪器而导致部分指标不能广泛应用. ...
The effect of high temperature stress on the phenology, growth and yield of five wheat (Triticum aestivum L.) genotypes
2
2012
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
Accelerated dryland expansion under climate change
2
2016
... 受全球气候变化和经济快速发展的影响,全球干旱呈现多发、加重趋势,中亚地区65%的区域表现为干旱程度加剧、强度持续增强,2022年全球极端干旱问题尤为突出(李稚等,2022;张强,2022;Metz et al.,2007;Li et al.,2015;IPCC,2022).随着全球气候变暖,水循环加快,陆地蒸散发和储水量减少,干旱半干旱区不断扩张,干旱持续加剧,干旱风险进一步加大,且存在较大的区域差异(张强等,2017;Salinger et al.,2005;Cook et al.,2007;Metz et al.,2007;Huang et al.,2016).联合国《2022年全球干旱数字》报告指出,自2000年以来全球干旱次数增加了29%,全球处于高温和干旱管理的“十字路口”(UNCCD, 2022).预计到2100年,极端干旱地区的干旱次数和强度将增加1%~30%(IPCC,2007). ...
... 我国是一个农业大国,也是旱灾频发的国家,旱灾影响范围广、历时长、危害重.据统计,我国每年因干旱受灾的农田面积多达2 600万hm2,粮食减产高达50%~75%(姚玉璧等,2013;张强等,2014).近年来,受气候变化和水资源短缺的影响,我国农作物受旱、成灾和绝收面积不断扩大,在北方干旱形势依然严峻的情况下南方干旱出现明显增加和加重趋势,新旱区逐渐扩展,局部性、区域性干旱灾害频频出现,损失巨大(张强等,2014;康蕾和张红旗,2014;倪深海等,2022;张强,2022),严重威胁着国家的粮食安全和生态安全(王春乙,2007;张强等,2012a;张强等,2012b;肖国举和李裕,2012;康蕾和张红旗,2014;周广胜等,2016;Fu et al.,2013),已成为制约社会经济可持续发展的最主要因素,给我国农业生产和抗旱减灾工作带来前所未有的挑战和困难(吕娟等,2011;张强等,2014;Huang et al.,2016). ...
2
2007
... 受全球气候变化和经济快速发展的影响,全球干旱呈现多发、加重趋势,中亚地区65%的区域表现为干旱程度加剧、强度持续增强,2022年全球极端干旱问题尤为突出(李稚等,2022;张强,2022;Metz et al.,2007;Li et al.,2015;IPCC,2022).随着全球气候变暖,水循环加快,陆地蒸散发和储水量减少,干旱半干旱区不断扩张,干旱持续加剧,干旱风险进一步加大,且存在较大的区域差异(张强等,2017;Salinger et al.,2005;Cook et al.,2007;Metz et al.,2007;Huang et al.,2016).联合国《2022年全球干旱数字》报告指出,自2000年以来全球干旱次数增加了29%,全球处于高温和干旱管理的“十字路口”(UNCCD, 2022).预计到2100年,极端干旱地区的干旱次数和强度将增加1%~30%(IPCC,2007). ...
... 农业干旱是农作物生长发育过程中,因降雨不足或长期无雨造成大气干旱、土壤缺水,致使作物得不到适时适量的水分补给,无法满足正常需水,影响其光合过程及生物量积累,最终造成农作物减产甚至绝收.农业干旱是影响农业生产的重要因素,也是农业生产面临的最大风险之一(王春乙,2007;肖国举和李裕,2012;张强等,2017;Mirzabaev et al.,2023).农业生产决定着粮食产量的稳定,是国家战略性、基础性核心产业,农作物种植是农业的“芯片”,是促进农业长期稳定发展,保障国家粮食安全的根本.干旱缺水直接影响农作物的生长发育和产量形成,造成粮食产量下降、品质降低,从而增加粮食生产供给的不稳定性和风险,直接威胁粮食安全,成为制约社会经济发展的重要因素之一(姚玉璧等,2013;张强等,2008;张强等,2020;IPCC,2007;IPCC,2022;Mirzabaev et al.,2023),长期以来农业旱灾对粮食安全的影响是学者和政府部门关注的重点和焦点.农业干旱对作物的影响从微观到宏观表现在细胞、生理和形态等不同层次水平上,其影响程度取决于干旱开始时间、发展和持续时间、严重程度及农作物种类和品种、所处生育期等因素(赵鸿等,2016;Ulrich et al.,2019;Gupta et al.,2020). ...
Climate Change 2022: Impacts, Adaptation and Vulnerability
4
2023
... 受全球气候变化和经济快速发展的影响,全球干旱呈现多发、加重趋势,中亚地区65%的区域表现为干旱程度加剧、强度持续增强,2022年全球极端干旱问题尤为突出(李稚等,2022;张强,2022;Metz et al.,2007;Li et al.,2015;IPCC,2022).随着全球气候变暖,水循环加快,陆地蒸散发和储水量减少,干旱半干旱区不断扩张,干旱持续加剧,干旱风险进一步加大,且存在较大的区域差异(张强等,2017;Salinger et al.,2005;Cook et al.,2007;Metz et al.,2007;Huang et al.,2016).联合国《2022年全球干旱数字》报告指出,自2000年以来全球干旱次数增加了29%,全球处于高温和干旱管理的“十字路口”(UNCCD, 2022).预计到2100年,极端干旱地区的干旱次数和强度将增加1%~30%(IPCC,2007). ...
... 农业干旱是农作物生长发育过程中,因降雨不足或长期无雨造成大气干旱、土壤缺水,致使作物得不到适时适量的水分补给,无法满足正常需水,影响其光合过程及生物量积累,最终造成农作物减产甚至绝收.农业干旱是影响农业生产的重要因素,也是农业生产面临的最大风险之一(王春乙,2007;肖国举和李裕,2012;张强等,2017;Mirzabaev et al.,2023).农业生产决定着粮食产量的稳定,是国家战略性、基础性核心产业,农作物种植是农业的“芯片”,是促进农业长期稳定发展,保障国家粮食安全的根本.干旱缺水直接影响农作物的生长发育和产量形成,造成粮食产量下降、品质降低,从而增加粮食生产供给的不稳定性和风险,直接威胁粮食安全,成为制约社会经济发展的重要因素之一(姚玉璧等,2013;张强等,2008;张强等,2020;IPCC,2007;IPCC,2022;Mirzabaev et al.,2023),长期以来农业旱灾对粮食安全的影响是学者和政府部门关注的重点和焦点.农业干旱对作物的影响从微观到宏观表现在细胞、生理和形态等不同层次水平上,其影响程度取决于干旱开始时间、发展和持续时间、严重程度及农作物种类和品种、所处生育期等因素(赵鸿等,2016;Ulrich et al.,2019;Gupta et al.,2020). ...
... 联合国《2022年干旱数字》报告指出,干旱对世界农业生产产生了深远、广泛且低估了的影响,尤其对粮食安全的影响(Gupta et al.,2020;Mirzabaev et al.,2023).干旱引起的水分短缺会对农作物造成一定危害,导致作物产量降低、品质下降,甚至绝收.IPCC第六次评估报告(IPCC,2022)指出,21世纪气候变化对粮食产量的影响仍以负面为主,变幅随不同作物、时间、地区变化差异较大.气候变化使玉米、大豆、水稻、小麦产量每10 a分别减少2.3%、3.3%、0.7%、1.3%,据统计,每年因干旱造成的粮食减产超过其他因素造成的总和,在过去10 a中全球因干旱造成的作物产量损失约300亿美元(FAO,2018).图1展示了全球人口、可耕地面积、农业淡水需求量的过去、现在和未来(Gupta et al.,2020).可以看出,全球人口从1990年的50亿增加到目前的75亿多,预计到2050年将增加到97~100亿[图1(a)],同时全球粮食需求量也将加速增长,从如今的约21亿t增长到2050年的30亿t左右,届时预计有50亿人生活在缺水地区(FAO,2006;Gupta et al.,2020);尽管全球可耕地面积略有增加[图1(b)],考虑到目前每公顷作物产量的比率,预计需要额外增加100万hm2可耕土地才能确保粮食安全(Gupta et al.,2020);考虑到目前气候变化趋势,预计到2050年农业用水需求量可能增加1倍[图1(c)],淡水供应量将下降50%(Gleick,2000;Koncagül et al.,2020;Gupta et al.,2020). ...
... 图6是1980年以来全国9大地区重旱以上灾害发生频率.可以看出,1980年以后我国北方的内蒙古、西北地区、东北地区和黄淮海地区干旱灾害发生频繁且较为严重,重旱以上发生频率分别为45.0%、40.0%、35.0%和25.0%,受灾率和成灾率均超过15.0%和10.0%(倪深海等,2022).从干旱发生范围来看,1980年以前发生重旱以上的区域波及10个省(市、区),而1980年以后扩增到16个省(市、区)(倪深海等,2022).受气候变暖影响,近50 a来西北地区干旱频率、强度和受灾面积增加,损失加重.其中,春旱、秋旱频次增加,夏旱频次减少,春旱、秋旱多于夏旱,特、重旱多出现在春季,主要发生在西北地区东部,尤其是甘肃河东地区,如1995年干旱最严重,受灾面积208.7万hm2,成灾面积170.8万hm2,成灾率为45%,粮食减产150万t;其次是2000年,成灾率达34.9%,粮食减产135万t(邓振镛等,2007).预计未来一段时间内,如果不采取有效应对措施,在气候变暖背景下到2030年中国种植业生产力总体可能下降5%~10%,其中小麦、水稻和玉米三大作物均以降低为主,2050年后受影响会更大(IPCC,2022). ...
Responses of potato genotypes to drought. I. Expansion of individual leaves and osmotic adjustment
2
1993
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 在农业干旱监测中,农业干旱指标主要有气象指标、土壤墒情指标、作物生理生态指标和一些其他指标.气象指标包括降水距平百分数、连续无有效降水日数、湿度指标和Z指数等(孙荣强,1994;姚玉璧等,2013;国家气象中心等,2015;Zhang et al.,2009).土壤墒情指标包括土壤相对湿度、土壤有效水分存储量、土壤水分旱灾指数等(国家气象中心等,2015;Hollinger et al.,1993;Dai et al.,2004).作物生理生态指标主要是一些基于植株生长和光合特性的生理参数,用来衡量植株水分亏缺状况,如光合速率、蒸腾速率、气孔导度、复水后的光合恢复、叶水势、茎秆水势、叶绿素、叶片相对含水量、叶片扩展速率、作物水分胁迫指数、冠层温度、块茎碳同位素鉴别等(姚玉璧等,2013;张强等,2014;赵鸿等,2018;纪瑞鹏等,2019;Jefferies,1993;Medrano et al.,2002;Zakaluk and Ranjan,2006;Ricardo,2012;Ramirez et al.,2016),这些基于植株的监测指标更加方便、适时,但有些地区由于没有专用测量仪器而导致部分指标不能广泛应用. ...
Current topics in drought physiology
1
1992
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
Rate of development of postanthesis water deficits and grain filling of spring wheat
1
1992
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
1
2020
... 联合国《2022年干旱数字》报告指出,干旱对世界农业生产产生了深远、广泛且低估了的影响,尤其对粮食安全的影响(Gupta et al.,2020;Mirzabaev et al.,2023).干旱引起的水分短缺会对农作物造成一定危害,导致作物产量降低、品质下降,甚至绝收.IPCC第六次评估报告(IPCC,2022)指出,21世纪气候变化对粮食产量的影响仍以负面为主,变幅随不同作物、时间、地区变化差异较大.气候变化使玉米、大豆、水稻、小麦产量每10 a分别减少2.3%、3.3%、0.7%、1.3%,据统计,每年因干旱造成的粮食减产超过其他因素造成的总和,在过去10 a中全球因干旱造成的作物产量损失约300亿美元(FAO,2018).图1展示了全球人口、可耕地面积、农业淡水需求量的过去、现在和未来(Gupta et al.,2020).可以看出,全球人口从1990年的50亿增加到目前的75亿多,预计到2050年将增加到97~100亿[图1(a)],同时全球粮食需求量也将加速增长,从如今的约21亿t增长到2050年的30亿t左右,届时预计有50亿人生活在缺水地区(FAO,2006;Gupta et al.,2020);尽管全球可耕地面积略有增加[图1(b)],考虑到目前每公顷作物产量的比率,预计需要额外增加100万hm2可耕土地才能确保粮食安全(Gupta et al.,2020);考虑到目前气候变化趋势,预计到2050年农业用水需求量可能增加1倍[图1(c)],淡水供应量将下降50%(Gleick,2000;Koncagül et al.,2020;Gupta et al.,2020). ...
Influence of extreme weather disasters on global crop production
2
2016
... 2022年气候异常,北半球遭遇了大范围、史无前例的严重干旱,北美洲、欧洲、地中海地区、东北非地区以及中国南方地区都出现破纪录的极端酷热天气,许多地区经历了极端干旱(图2),对农业生产产生较大影响,大部分农作物有一定程度的减产.2022年9月联合国粮农组织发布的月报,将2022年全球谷物产量预测从7月初预测的27.92亿t下调至27.74亿t,比2021年预估产量低1.4%(https://xueqiu.com/4587623715/231014661).1964—2007年极端天气灾害造成全球粮食作物大面积减产,尤其是玉米、水稻、小麦,其中干旱和极端高温导致各国粮食减产9%~10%,发达国家的粮食产量损失比发展中国家高出8%~11%(纪瑞鹏等,2019;Lesk et al.,2016;Mirzabaev et al.,2023) ...
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
Exploration of copula function use in crop meteorological drought risk analysis: a case study of winter wheat in Beijing, China
2
2015
... 干旱灾害(简称“旱灾”),是指一段时间内降水量比常年明显偏少,造成某一地区经济社会活动(尤其是农业生产)和人类生活遭受较大危害的现象,普遍存在于世界各地,频繁发生于不同时期(Wilhite,2000;Li et al.,2015).旱灾是气象干旱、农业干旱、水文干旱、社会经济干旱和生态干旱之间的一种链状传递和转化过程,一个完整的干旱事件包括干旱历时、严重程度和平均强度(姚玉璧等,2013;张强等,2014;Heffernan,2013).全球每年因干旱导致的经济损失达60~80亿美元,2021年甚至高达121亿美元(American Meteorological Society, 1997;应急管理部-教育部减灾与应急管理研究院等,2022),远高于其他气象灾害造成的损失(张强等,2014;Wilhite,2000),受到高度关注(王春乙,2007;吕娟等,2011;Turner et al., 2011;Gupta et al.,2020). ...
... 受全球气候变化和经济快速发展的影响,全球干旱呈现多发、加重趋势,中亚地区65%的区域表现为干旱程度加剧、强度持续增强,2022年全球极端干旱问题尤为突出(李稚等,2022;张强,2022;Metz et al.,2007;Li et al.,2015;IPCC,2022).随着全球气候变暖,水循环加快,陆地蒸散发和储水量减少,干旱半干旱区不断扩张,干旱持续加剧,干旱风险进一步加大,且存在较大的区域差异(张强等,2017;Salinger et al.,2005;Cook et al.,2007;Metz et al.,2007;Huang et al.,2016).联合国《2022年全球干旱数字》报告指出,自2000年以来全球干旱次数增加了29%,全球处于高温和干旱管理的“十字路口”(UNCCD, 2022).预计到2100年,极端干旱地区的干旱次数和强度将增加1%~30%(IPCC,2007). ...
Growth and phenology of three Andean potato varieties (Solanum tuberosum L.) under water stress
1
2016
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
Combined effects of drought and high temperature on water relations of wheat and sorghum
2
2001
... 从分子生物学角度看,细胞水分损失是干旱胁迫的标志(Gupta et al.,2020).作物感知缺水信号并启动应对策略的能力被认为抗旱性.作物抗旱性是一个复杂性状,通常以避免或忍耐细胞脱水而防止水分流失来渡过干旱(Turner,1986;Machado and Paulsen,2001),主要表现为逃旱、避旱和耐旱等.逃旱是指作物通过调控生长发育期(提前或滞后水分敏感时段)来逃避干旱胁迫,一般发生在作物生长受阻之前;避旱,为减少水分损失,在干旱条件下保持高水势的能力,作物通过增加体内含水量避免组织损伤,涉及植株快速生长、叶片卷曲、叶面积减小、气孔开闭等(Morgan,1984;Turner,1986;Morgan,1991;Machado and Paulsen,2001;Gupta et al.,2020);耐旱是指组织内部含水量较低,干旱期间仍需保持生长耐力,在水分亏缺加重情况下通过保持充足的细胞膨胀来维持新陈代谢(Gupta et al.,2020). ...
... ;Machado and Paulsen,2001;Gupta et al.,2020);耐旱是指组织内部含水量较低,干旱期间仍需保持生长耐力,在水分亏缺加重情况下通过保持充足的细胞膨胀来维持新陈代谢(Gupta et al.,2020). ...
The influence of early soil moisture stress on tuber numbers in potato
1
1986
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars
1
2010
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
Molecular and physiological adaptation to prolonged drought stress in the leaves of two Andean potato genotypes
1
2008
... 植物生长调节剂(外用)和植物激素(内用)是影响植物生理过程的物质,其浓度很低时起调节作用,如脱落酸、乙烯、生长素、赤霉素、细胞分裂素等(Morgan,1991;康书江等,1997).在分子和生化水平上,植物通过在转录和蛋白表达过程中改变调节反应来感知和响应干旱胁迫,从而影响生化途径和代谢过程,进而影响生理和发育过程(Mane et al.,2008;Vasquez-Robinet et al.,2008;Zinselmeier et al.,1995).在土壤干旱条件下植物通常会产生脱落酸(Abscisic Acid,ABA),它是一种主要的化学根-芽胁迫信号(Davies and Zhang,1991),是通过激活对干旱响应的基因表达而表征细胞反应的一个关键信号(Chaves et al.,2003).ABA含量可作为植物抗旱性的评价指标之一,其与籽粒的生长速率有关.当土壤干旱时失水的根系产生根源信号ABA,通过木质部输送到地上部调节气孔开闭(Davies and Zhang,1991),作物根冠、叶片、花、籽粒生长进程中ABA含量明显上升(赵鸿等,2008;张玉书等,2012;Obidiegwu et al,2015),且随着干旱胁迫程度的加剧而增加,如受到水分胁迫时小麦籽粒生长减慢、结实率降低,玉米胚乳细胞分化率降低.乙烯(Ethylene)也是干旱胁迫下植物体产生的激素,在干旱感知时乙烯通过蒸腾流将其他激素信号与ABA一起发送到嫩芽中.干旱引发的乙烯释放过量,会引起小麦、玉米等籽粒产量下降(赵鸿等,2008;Obidiegwu et al.,2015). ...
The national and international importance of drought and salinity effects on agricultural production
2
1986
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
Regulation of photosynthesis of C3 plants in response to progressive drought: stomatal conductance as a reference parameter
2
2002
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 在农业干旱监测中,农业干旱指标主要有气象指标、土壤墒情指标、作物生理生态指标和一些其他指标.气象指标包括降水距平百分数、连续无有效降水日数、湿度指标和Z指数等(孙荣强,1994;姚玉璧等,2013;国家气象中心等,2015;Zhang et al.,2009).土壤墒情指标包括土壤相对湿度、土壤有效水分存储量、土壤水分旱灾指数等(国家气象中心等,2015;Hollinger et al.,1993;Dai et al.,2004).作物生理生态指标主要是一些基于植株生长和光合特性的生理参数,用来衡量植株水分亏缺状况,如光合速率、蒸腾速率、气孔导度、复水后的光合恢复、叶水势、茎秆水势、叶绿素、叶片相对含水量、叶片扩展速率、作物水分胁迫指数、冠层温度、块茎碳同位素鉴别等(姚玉璧等,2013;张强等,2014;赵鸿等,2018;纪瑞鹏等,2019;Jefferies,1993;Medrano et al.,2002;Zakaluk and Ranjan,2006;Ricardo,2012;Ramirez et al.,2016),这些基于植株的监测指标更加方便、适时,但有些地区由于没有专用测量仪器而导致部分指标不能广泛应用. ...
2
2007
... 受全球气候变化和经济快速发展的影响,全球干旱呈现多发、加重趋势,中亚地区65%的区域表现为干旱程度加剧、强度持续增强,2022年全球极端干旱问题尤为突出(李稚等,2022;张强,2022;Metz et al.,2007;Li et al.,2015;IPCC,2022).随着全球气候变暖,水循环加快,陆地蒸散发和储水量减少,干旱半干旱区不断扩张,干旱持续加剧,干旱风险进一步加大,且存在较大的区域差异(张强等,2017;Salinger et al.,2005;Cook et al.,2007;Metz et al.,2007;Huang et al.,2016).联合国《2022年全球干旱数字》报告指出,自2000年以来全球干旱次数增加了29%,全球处于高温和干旱管理的“十字路口”(UNCCD, 2022).预计到2100年,极端干旱地区的干旱次数和强度将增加1%~30%(IPCC,2007). ...
... ;Metz et al.,2007;Huang et al.,2016).联合国《2022年全球干旱数字》报告指出,自2000年以来全球干旱次数增加了29%,全球处于高温和干旱管理的“十字路口”(UNCCD, 2022).预计到2100年,极端干旱地区的干旱次数和强度将增加1%~30%(IPCC,2007). ...
Severe climate change risks to food security and nutrition
4
2023
... 农业干旱是农作物生长发育过程中,因降雨不足或长期无雨造成大气干旱、土壤缺水,致使作物得不到适时适量的水分补给,无法满足正常需水,影响其光合过程及生物量积累,最终造成农作物减产甚至绝收.农业干旱是影响农业生产的重要因素,也是农业生产面临的最大风险之一(王春乙,2007;肖国举和李裕,2012;张强等,2017;Mirzabaev et al.,2023).农业生产决定着粮食产量的稳定,是国家战略性、基础性核心产业,农作物种植是农业的“芯片”,是促进农业长期稳定发展,保障国家粮食安全的根本.干旱缺水直接影响农作物的生长发育和产量形成,造成粮食产量下降、品质降低,从而增加粮食生产供给的不稳定性和风险,直接威胁粮食安全,成为制约社会经济发展的重要因素之一(姚玉璧等,2013;张强等,2008;张强等,2020;IPCC,2007;IPCC,2022;Mirzabaev et al.,2023),长期以来农业旱灾对粮食安全的影响是学者和政府部门关注的重点和焦点.农业干旱对作物的影响从微观到宏观表现在细胞、生理和形态等不同层次水平上,其影响程度取决于干旱开始时间、发展和持续时间、严重程度及农作物种类和品种、所处生育期等因素(赵鸿等,2016;Ulrich et al.,2019;Gupta et al.,2020). ...
... ;Mirzabaev et al.,2023),长期以来农业旱灾对粮食安全的影响是学者和政府部门关注的重点和焦点.农业干旱对作物的影响从微观到宏观表现在细胞、生理和形态等不同层次水平上,其影响程度取决于干旱开始时间、发展和持续时间、严重程度及农作物种类和品种、所处生育期等因素(赵鸿等,2016;Ulrich et al.,2019;Gupta et al.,2020). ...
... 联合国《2022年干旱数字》报告指出,干旱对世界农业生产产生了深远、广泛且低估了的影响,尤其对粮食安全的影响(Gupta et al.,2020;Mirzabaev et al.,2023).干旱引起的水分短缺会对农作物造成一定危害,导致作物产量降低、品质下降,甚至绝收.IPCC第六次评估报告(IPCC,2022)指出,21世纪气候变化对粮食产量的影响仍以负面为主,变幅随不同作物、时间、地区变化差异较大.气候变化使玉米、大豆、水稻、小麦产量每10 a分别减少2.3%、3.3%、0.7%、1.3%,据统计,每年因干旱造成的粮食减产超过其他因素造成的总和,在过去10 a中全球因干旱造成的作物产量损失约300亿美元(FAO,2018).图1展示了全球人口、可耕地面积、农业淡水需求量的过去、现在和未来(Gupta et al.,2020).可以看出,全球人口从1990年的50亿增加到目前的75亿多,预计到2050年将增加到97~100亿[图1(a)],同时全球粮食需求量也将加速增长,从如今的约21亿t增长到2050年的30亿t左右,届时预计有50亿人生活在缺水地区(FAO,2006;Gupta et al.,2020);尽管全球可耕地面积略有增加[图1(b)],考虑到目前每公顷作物产量的比率,预计需要额外增加100万hm2可耕土地才能确保粮食安全(Gupta et al.,2020);考虑到目前气候变化趋势,预计到2050年农业用水需求量可能增加1倍[图1(c)],淡水供应量将下降50%(Gleick,2000;Koncagül et al.,2020;Gupta et al.,2020). ...
... 2022年气候异常,北半球遭遇了大范围、史无前例的严重干旱,北美洲、欧洲、地中海地区、东北非地区以及中国南方地区都出现破纪录的极端酷热天气,许多地区经历了极端干旱(图2),对农业生产产生较大影响,大部分农作物有一定程度的减产.2022年9月联合国粮农组织发布的月报,将2022年全球谷物产量预测从7月初预测的27.92亿t下调至27.74亿t,比2021年预估产量低1.4%(https://xueqiu.com/4587623715/231014661).1964—2007年极端天气灾害造成全球粮食作物大面积减产,尤其是玉米、水稻、小麦,其中干旱和极端高温导致各国粮食减产9%~10%,发达国家的粮食产量损失比发展中国家高出8%~11%(纪瑞鹏等,2019;Lesk et al.,2016;Mirzabaev et al.,2023) ...
Osmoregulation and water stress in higher plants
1
1984
... 从分子生物学角度看,细胞水分损失是干旱胁迫的标志(Gupta et al.,2020).作物感知缺水信号并启动应对策略的能力被认为抗旱性.作物抗旱性是一个复杂性状,通常以避免或忍耐细胞脱水而防止水分流失来渡过干旱(Turner,1986;Machado and Paulsen,2001),主要表现为逃旱、避旱和耐旱等.逃旱是指作物通过调控生长发育期(提前或滞后水分敏感时段)来逃避干旱胁迫,一般发生在作物生长受阻之前;避旱,为减少水分损失,在干旱条件下保持高水势的能力,作物通过增加体内含水量避免组织损伤,涉及植株快速生长、叶片卷曲、叶面积减小、气孔开闭等(Morgan,1984;Turner,1986;Morgan,1991;Machado and Paulsen,2001;Gupta et al.,2020);耐旱是指组织内部含水量较低,干旱期间仍需保持生长耐力,在水分亏缺加重情况下通过保持充足的细胞膨胀来维持新陈代谢(Gupta et al.,2020). ...
Effects of abiotic stresses on plant hormone systems
2
1991
... 植物生长调节剂(外用)和植物激素(内用)是影响植物生理过程的物质,其浓度很低时起调节作用,如脱落酸、乙烯、生长素、赤霉素、细胞分裂素等(Morgan,1991;康书江等,1997).在分子和生化水平上,植物通过在转录和蛋白表达过程中改变调节反应来感知和响应干旱胁迫,从而影响生化途径和代谢过程,进而影响生理和发育过程(Mane et al.,2008;Vasquez-Robinet et al.,2008;Zinselmeier et al.,1995).在土壤干旱条件下植物通常会产生脱落酸(Abscisic Acid,ABA),它是一种主要的化学根-芽胁迫信号(Davies and Zhang,1991),是通过激活对干旱响应的基因表达而表征细胞反应的一个关键信号(Chaves et al.,2003).ABA含量可作为植物抗旱性的评价指标之一,其与籽粒的生长速率有关.当土壤干旱时失水的根系产生根源信号ABA,通过木质部输送到地上部调节气孔开闭(Davies and Zhang,1991),作物根冠、叶片、花、籽粒生长进程中ABA含量明显上升(赵鸿等,2008;张玉书等,2012;Obidiegwu et al,2015),且随着干旱胁迫程度的加剧而增加,如受到水分胁迫时小麦籽粒生长减慢、结实率降低,玉米胚乳细胞分化率降低.乙烯(Ethylene)也是干旱胁迫下植物体产生的激素,在干旱感知时乙烯通过蒸腾流将其他激素信号与ABA一起发送到嫩芽中.干旱引发的乙烯释放过量,会引起小麦、玉米等籽粒产量下降(赵鸿等,2008;Obidiegwu et al.,2015). ...
... 从分子生物学角度看,细胞水分损失是干旱胁迫的标志(Gupta et al.,2020).作物感知缺水信号并启动应对策略的能力被认为抗旱性.作物抗旱性是一个复杂性状,通常以避免或忍耐细胞脱水而防止水分流失来渡过干旱(Turner,1986;Machado and Paulsen,2001),主要表现为逃旱、避旱和耐旱等.逃旱是指作物通过调控生长发育期(提前或滞后水分敏感时段)来逃避干旱胁迫,一般发生在作物生长受阻之前;避旱,为减少水分损失,在干旱条件下保持高水势的能力,作物通过增加体内含水量避免组织损伤,涉及植株快速生长、叶片卷曲、叶面积减小、气孔开闭等(Morgan,1984;Turner,1986;Morgan,1991;Machado and Paulsen,2001;Gupta et al.,2020);耐旱是指组织内部含水量较低,干旱期间仍需保持生长耐力,在水分亏缺加重情况下通过保持充足的细胞膨胀来维持新陈代谢(Gupta et al.,2020). ...
Effect of drought on metabolism and partitioning of carbon in two wheat varieties differing in drought-tolerance
1
1985
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
Coping with drought: stress and adaptive responses in potato and perspectives for improvement
6
2015
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
... 植物生长调节剂(外用)和植物激素(内用)是影响植物生理过程的物质,其浓度很低时起调节作用,如脱落酸、乙烯、生长素、赤霉素、细胞分裂素等(Morgan,1991;康书江等,1997).在分子和生化水平上,植物通过在转录和蛋白表达过程中改变调节反应来感知和响应干旱胁迫,从而影响生化途径和代谢过程,进而影响生理和发育过程(Mane et al.,2008;Vasquez-Robinet et al.,2008;Zinselmeier et al.,1995).在土壤干旱条件下植物通常会产生脱落酸(Abscisic Acid,ABA),它是一种主要的化学根-芽胁迫信号(Davies and Zhang,1991),是通过激活对干旱响应的基因表达而表征细胞反应的一个关键信号(Chaves et al.,2003).ABA含量可作为植物抗旱性的评价指标之一,其与籽粒的生长速率有关.当土壤干旱时失水的根系产生根源信号ABA,通过木质部输送到地上部调节气孔开闭(Davies and Zhang,1991),作物根冠、叶片、花、籽粒生长进程中ABA含量明显上升(赵鸿等,2008;张玉书等,2012;Obidiegwu et al,2015),且随着干旱胁迫程度的加剧而增加,如受到水分胁迫时小麦籽粒生长减慢、结实率降低,玉米胚乳细胞分化率降低.乙烯(Ethylene)也是干旱胁迫下植物体产生的激素,在干旱感知时乙烯通过蒸腾流将其他激素信号与ABA一起发送到嫩芽中.干旱引发的乙烯释放过量,会引起小麦、玉米等籽粒产量下降(赵鸿等,2008;Obidiegwu et al.,2015). ...
... ;Obidiegwu et al.,2015). ...
... 叶片对CO2的同化主要受气孔关闭、膜损伤和各种酶活性的干扰,尤其是CO2固定酶和三磷酸腺苷(ATP)合成酶,通过光呼吸途径增强的代谢物通量增加组织的氧化负荷,该过程产生活性氧,干旱胁迫下活性氧对生物大分子的损伤是影响植物生长的主要因素之一(Tezara et al.,1999;Farooq et al.,2009).干旱缺水时,作物体内活性氧的积累受到破坏,抗氧化酶活性降低,对活性氧的清除能力下降,致使作物避免或减轻受氧化伤害(赵鸿等,2008;Obidiegwu et al.,2015).轻度干旱可引起叶片中过氧化氢酶(Catalase,CAT)活性升高,而重度干旱下CAT活性降低.随着干旱胁迫的持续,叶片中过氧化物酶(Peroxidase,POD)活性渐增,丙二醛(Malonic Dialdehyde,MDA)含量增大,膜脂、蛋白质和核酸氧化加剧,明显抑制了植株的生长发育(张玉书等,2012;Obidiegwu et al.,2015).马铃薯在遭遇不同程度干旱胁迫时,随着胁迫程度的增加,植株体内超氧化物歧化酶(Superoxide Dismutase,SOD)活性下降,马铃薯受自由基损伤加重,无法起到保护细胞、去除活性氧自由基的作用,细胞受伤害增大、植株老化加速.在极度干旱胁迫下,当活性氧浓度超出清除剂的潜力时,干旱对不同植物细胞包括脂质、蛋白质和脱氧核糖核酸(Deoxyribo Nucleic Acid,DNA)造成不可修复的损害,导致植物细胞死亡(Obidiegwu et al.,2015;Gupta et al.,2020). ...
... ;Obidiegwu et al.,2015).马铃薯在遭遇不同程度干旱胁迫时,随着胁迫程度的增加,植株体内超氧化物歧化酶(Superoxide Dismutase,SOD)活性下降,马铃薯受自由基损伤加重,无法起到保护细胞、去除活性氧自由基的作用,细胞受伤害增大、植株老化加速.在极度干旱胁迫下,当活性氧浓度超出清除剂的潜力时,干旱对不同植物细胞包括脂质、蛋白质和脱氧核糖核酸(Deoxyribo Nucleic Acid,DNA)造成不可修复的损害,导致植物细胞死亡(Obidiegwu et al.,2015;Gupta et al.,2020). ...
... ).马铃薯在遭遇不同程度干旱胁迫时,随着胁迫程度的增加,植株体内超氧化物歧化酶(Superoxide Dismutase,SOD)活性下降,马铃薯受自由基损伤加重,无法起到保护细胞、去除活性氧自由基的作用,细胞受伤害增大、植株老化加速.在极度干旱胁迫下,当活性氧浓度超出清除剂的潜力时,干旱对不同植物细胞包括脂质、蛋白质和脱氧核糖核酸(Deoxyribo Nucleic Acid,DNA)造成不可修复的损害,导致植物细胞死亡(Obidiegwu et al.,2015;Gupta et al.,2020). ...
Keeping track of crop moisture conditions, nationwide: the new crop moisture index
2
1968
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 此外,还有一些重要的监测指标如帕尔默干旱指数、作物水分亏缺指数、作物水分指数、农田与作物形态农业干旱指标、作物特征旱灾指数、减产率农业干旱指数、综合农业干旱指数、区域性农业干旱指标以及农业干旱过程的确定和评估等(Palmer,1968;李柏贞和周广胜,2014;国家气象中心等,2015).其中,帕尔默干旱指数是目前国际上应用最广泛的干旱指标,它引入了水量平衡概念,包含降水量、蒸散量、径流量和土壤水分储存量等,用于监测以月为时间尺度的干旱(表1).该指数不仅考虑了当前的水分条件,还考虑了前期水分状况及干旱持续时间等,具有较好的时空可比性,能够较好地监测评估较长时期的干旱,同时也是衡量土壤湿度和确定干旱开始和终止时间最有效的评估方法,但该指标考虑的因素较多,对资料要求较高,无法实现作物对水分亏缺响应的逐日监测. ...
Advances in Land surface models and indicators for drought monitoring and prediction
1
2021
... 美国、澳大利亚等受干旱影响较重的国家,在过去30 a间,率先摒弃了被动应急抗旱的方式,积极采取主动的干旱风险管理模式,即预警防旱早于应急救援(Collins et al.,2016),有效应对可能发生的干旱灾害(Fu et al.,2013;Hervás-Gámez and Delgado-Ramos,2020).具体包括3个方面:首先,通过制定抗旱预案制度、发布国家和区域干旱政策、提高干旱监测预警能力、加强干旱灾害风险评估等手段,增强全社会防御干旱的能力(Wilhite et al.,2007;Peters-lidard et al.,2021;Steinemann et al.,2015;Zarei et al.,2020);其次,在干旱易发区域或干旱发生后,采取强化各部门需水管理、普及淡水资源化以及提高水分利用效率措施,如种植抗旱作物、农业节水灌溉、废水再利用及微咸水灌溉等,也可采用工程抗旱、生物抗旱等措施尽可能降低旱灾造成的损失(WMO and GWP,2014);第三,对于干旱造成损失的,政府制定相应政策评估受损,给予救济金补贴,让受灾严重的群体尽快恢复生产生活(WMO and GWP,2017).此外,澳大利亚还利用地下含水层储水-回灌技术发展农业节水灌溉,即把冬天地表多余的水资源通过压力储存到地下100 m左右的含水层,遇到干旱月份再抽出来进行农田灌溉;通过地理信息系统(Geographic Information System,GIS)、遥感系统(Remote Sensing,RS)、全球定位系统(Global Position System,GPS)等3S系统和制图(Mapping)、监控(Monitoring)、管理(Management)系统等3M系统进行节水灌溉,以实现节约水资源的目的(https://world.qianlong.com/2022/0830/7575142.shtml). ...
Photosynthesis and drought: can we make metabolic connections from available data?
2
2011
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
Maize yield, rainwater and nitrogen use efficiency as affected by maize genotypes and nitrogen rates on the Loess Plateau of China
2
2019a
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 第五,改进施肥方式和用量,测土施肥,配合深施、混施等施肥方式,提高肥效和作物对营养元素的利用率,如玉米施氮增效不仅可以获得较高产量、水分和氮素利用效率,还能降低硝态氮的残留量,有效减缓土壤污染(肖国举和李裕,2012;Qiang et al.,2019a).冬小麦测土施肥,可用于诊断其氮营养状况,提高水分利用效率、增加叶面积指数和干物质积累(肖国举和李裕,2012;Qiang et al.,2019b).未来半干旱地区的小麦生产,可视温度、CO2浓度升高情况适量采用低磷肥、低氮肥投入,这将更经济、适宜(王润元等,2015;肖国举和李裕,2012;鲍文中和周广胜,2017). ...
Determination of critical nitrogen dilution curve based on leaf area index for winter wheat in the Guanzhong Plain, Northwest China
1
2019b
... 第五,改进施肥方式和用量,测土施肥,配合深施、混施等施肥方式,提高肥效和作物对营养元素的利用率,如玉米施氮增效不仅可以获得较高产量、水分和氮素利用效率,还能降低硝态氮的残留量,有效减缓土壤污染(肖国举和李裕,2012;Qiang et al.,2019a).冬小麦测土施肥,可用于诊断其氮营养状况,提高水分利用效率、增加叶面积指数和干物质积累(肖国举和李裕,2012;Qiang et al.,2019b).未来半干旱地区的小麦生产,可视温度、CO2浓度升高情况适量采用低磷肥、低氮肥投入,这将更经济、适宜(王润元等,2015;肖国举和李裕,2012;鲍文中和周广胜,2017). ...
An efficient greenhouse method to screen potato genotypes for drought tolerance
1
2019
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
Defining biological thresholds associated to plant water status for monitoring water restriction effects: stomatal conductance and photosynthesis recovery as key indicators in potato
2
2016
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 在农业干旱监测中,农业干旱指标主要有气象指标、土壤墒情指标、作物生理生态指标和一些其他指标.气象指标包括降水距平百分数、连续无有效降水日数、湿度指标和Z指数等(孙荣强,1994;姚玉璧等,2013;国家气象中心等,2015;Zhang et al.,2009).土壤墒情指标包括土壤相对湿度、土壤有效水分存储量、土壤水分旱灾指数等(国家气象中心等,2015;Hollinger et al.,1993;Dai et al.,2004).作物生理生态指标主要是一些基于植株生长和光合特性的生理参数,用来衡量植株水分亏缺状况,如光合速率、蒸腾速率、气孔导度、复水后的光合恢复、叶水势、茎秆水势、叶绿素、叶片相对含水量、叶片扩展速率、作物水分胁迫指数、冠层温度、块茎碳同位素鉴别等(姚玉璧等,2013;张强等,2014;赵鸿等,2018;纪瑞鹏等,2019;Jefferies,1993;Medrano et al.,2002;Zakaluk and Ranjan,2006;Ricardo,2012;Ramirez et al.,2016),这些基于植株的监测指标更加方便、适时,但有些地区由于没有专用测量仪器而导致部分指标不能广泛应用. ...
Drought stress response in wheat: physiological and molecular analysis of resistant and sensitive genotypes
3
2006
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
Plant responses to drought stress from morphological to molecular features
4
2012
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
... ;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
... 在农业干旱监测中,农业干旱指标主要有气象指标、土壤墒情指标、作物生理生态指标和一些其他指标.气象指标包括降水距平百分数、连续无有效降水日数、湿度指标和Z指数等(孙荣强,1994;姚玉璧等,2013;国家气象中心等,2015;Zhang et al.,2009).土壤墒情指标包括土壤相对湿度、土壤有效水分存储量、土壤水分旱灾指数等(国家气象中心等,2015;Hollinger et al.,1993;Dai et al.,2004).作物生理生态指标主要是一些基于植株生长和光合特性的生理参数,用来衡量植株水分亏缺状况,如光合速率、蒸腾速率、气孔导度、复水后的光合恢复、叶水势、茎秆水势、叶绿素、叶片相对含水量、叶片扩展速率、作物水分胁迫指数、冠层温度、块茎碳同位素鉴别等(姚玉璧等,2013;张强等,2014;赵鸿等,2018;纪瑞鹏等,2019;Jefferies,1993;Medrano et al.,2002;Zakaluk and Ranjan,2006;Ricardo,2012;Ramirez et al.,2016),这些基于植株的监测指标更加方便、适时,但有些地区由于没有专用测量仪器而导致部分指标不能广泛应用. ...
Reducing vulnerability of agriculture and forestry to climate variability and change: workshop summary and recommendations
1
2005
... 受全球气候变化和经济快速发展的影响,全球干旱呈现多发、加重趋势,中亚地区65%的区域表现为干旱程度加剧、强度持续增强,2022年全球极端干旱问题尤为突出(李稚等,2022;张强,2022;Metz et al.,2007;Li et al.,2015;IPCC,2022).随着全球气候变暖,水循环加快,陆地蒸散发和储水量减少,干旱半干旱区不断扩张,干旱持续加剧,干旱风险进一步加大,且存在较大的区域差异(张强等,2017;Salinger et al.,2005;Cook et al.,2007;Metz et al.,2007;Huang et al.,2016).联合国《2022年全球干旱数字》报告指出,自2000年以来全球干旱次数增加了29%,全球处于高温和干旱管理的“十字路口”(UNCCD, 2022).预计到2100年,极端干旱地区的干旱次数和强度将增加1%~30%(IPCC,2007). ...
Variation in root traits of chickpea (Cicer arietinum L.) grown under terminal drought
1
2004
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
Developing and evaluating drought indicators for decision-making
1
2015
... 美国、澳大利亚等受干旱影响较重的国家,在过去30 a间,率先摒弃了被动应急抗旱的方式,积极采取主动的干旱风险管理模式,即预警防旱早于应急救援(Collins et al.,2016),有效应对可能发生的干旱灾害(Fu et al.,2013;Hervás-Gámez and Delgado-Ramos,2020).具体包括3个方面:首先,通过制定抗旱预案制度、发布国家和区域干旱政策、提高干旱监测预警能力、加强干旱灾害风险评估等手段,增强全社会防御干旱的能力(Wilhite et al.,2007;Peters-lidard et al.,2021;Steinemann et al.,2015;Zarei et al.,2020);其次,在干旱易发区域或干旱发生后,采取强化各部门需水管理、普及淡水资源化以及提高水分利用效率措施,如种植抗旱作物、农业节水灌溉、废水再利用及微咸水灌溉等,也可采用工程抗旱、生物抗旱等措施尽可能降低旱灾造成的损失(WMO and GWP,2014);第三,对于干旱造成损失的,政府制定相应政策评估受损,给予救济金补贴,让受灾严重的群体尽快恢复生产生活(WMO and GWP,2017).此外,澳大利亚还利用地下含水层储水-回灌技术发展农业节水灌溉,即把冬天地表多余的水资源通过压力储存到地下100 m左右的含水层,遇到干旱月份再抽出来进行农田灌溉;通过地理信息系统(Geographic Information System,GIS)、遥感系统(Remote Sensing,RS)、全球定位系统(Global Position System,GPS)等3S系统和制图(Mapping)、监控(Monitoring)、管理(Management)系统等3M系统进行节水灌溉,以实现节约水资源的目的(https://world.qianlong.com/2022/0830/7575142.shtml). ...
Plant adaptation to drought stress
2
2016
... 渗透调节、渗透保护、抗氧化和清除防御系统是作物抗旱性的重要基础.作物表现出一系列抵御干旱胁迫的机制,主要包括通过增加扩散阻力来减少水分流失、通过深层根系增强水分吸收和有效利用、通过更小和多汁的叶片来减少蒸腾损失(Farooq et al.,2009;Supratim et al.,2016).在营养物质中,钾离子有助于渗透调节.低分子渗透液,包括甜菜碱、脯氨酸和其他氨基酸、有机酸和多元醇等,在干旱条件下能维持细胞功能.在分子水平上目前已经鉴定出若干个干旱响应基因和转录因子,如脱水响应元件结合基因、水通道蛋白、胚胎发育后期丰富的蛋白质和脱氢蛋白等(Farooq et al.,2009;Supratim et al.,2016). ...
... ;Supratim et al.,2016). ...
Water stress inhibits plant photosynthesis by decreasing coupling factor and ATP
1
1999
... 叶片对CO2的同化主要受气孔关闭、膜损伤和各种酶活性的干扰,尤其是CO2固定酶和三磷酸腺苷(ATP)合成酶,通过光呼吸途径增强的代谢物通量增加组织的氧化负荷,该过程产生活性氧,干旱胁迫下活性氧对生物大分子的损伤是影响植物生长的主要因素之一(Tezara et al.,1999;Farooq et al.,2009).干旱缺水时,作物体内活性氧的积累受到破坏,抗氧化酶活性降低,对活性氧的清除能力下降,致使作物避免或减轻受氧化伤害(赵鸿等,2008;Obidiegwu et al.,2015).轻度干旱可引起叶片中过氧化氢酶(Catalase,CAT)活性升高,而重度干旱下CAT活性降低.随着干旱胁迫的持续,叶片中过氧化物酶(Peroxidase,POD)活性渐增,丙二醛(Malonic Dialdehyde,MDA)含量增大,膜脂、蛋白质和核酸氧化加剧,明显抑制了植株的生长发育(张玉书等,2012;Obidiegwu et al.,2015).马铃薯在遭遇不同程度干旱胁迫时,随着胁迫程度的增加,植株体内超氧化物歧化酶(Superoxide Dismutase,SOD)活性下降,马铃薯受自由基损伤加重,无法起到保护细胞、去除活性氧自由基的作用,细胞受伤害增大、植株老化加速.在极度干旱胁迫下,当活性氧浓度超出清除剂的潜力时,干旱对不同植物细胞包括脂质、蛋白质和脱氧核糖核酸(Deoxyribo Nucleic Acid,DNA)造成不可修复的损害,导致植物细胞死亡(Obidiegwu et al.,2015;Gupta et al.,2020). ...
Crop water deficits: a decade of progress
2
1986
... 从分子生物学角度看,细胞水分损失是干旱胁迫的标志(Gupta et al.,2020).作物感知缺水信号并启动应对策略的能力被认为抗旱性.作物抗旱性是一个复杂性状,通常以避免或忍耐细胞脱水而防止水分流失来渡过干旱(Turner,1986;Machado and Paulsen,2001),主要表现为逃旱、避旱和耐旱等.逃旱是指作物通过调控生长发育期(提前或滞后水分敏感时段)来逃避干旱胁迫,一般发生在作物生长受阻之前;避旱,为减少水分损失,在干旱条件下保持高水势的能力,作物通过增加体内含水量避免组织损伤,涉及植株快速生长、叶片卷曲、叶面积减小、气孔开闭等(Morgan,1984;Turner,1986;Morgan,1991;Machado and Paulsen,2001;Gupta et al.,2020);耐旱是指组织内部含水量较低,干旱期间仍需保持生长耐力,在水分亏缺加重情况下通过保持充足的细胞膨胀来维持新陈代谢(Gupta et al.,2020). ...
... ;Turner,1986;Morgan,1991;Machado and Paulsen,2001;Gupta et al.,2020);耐旱是指组织内部含水量较低,干旱期间仍需保持生长耐力,在水分亏缺加重情况下通过保持充足的细胞膨胀来维持新陈代谢(Gupta et al.,2020). ...
Climate change in south-west Australia and north-west China: challenges and opportunities for crop production
1
2011
... 干旱灾害(简称“旱灾”),是指一段时间内降水量比常年明显偏少,造成某一地区经济社会活动(尤其是农业生产)和人类生活遭受较大危害的现象,普遍存在于世界各地,频繁发生于不同时期(Wilhite,2000;Li et al.,2015).旱灾是气象干旱、农业干旱、水文干旱、社会经济干旱和生态干旱之间的一种链状传递和转化过程,一个完整的干旱事件包括干旱历时、严重程度和平均强度(姚玉璧等,2013;张强等,2014;Heffernan,2013).全球每年因干旱导致的经济损失达60~80亿美元,2021年甚至高达121亿美元(American Meteorological Society, 1997;应急管理部-教育部减灾与应急管理研究院等,2022),远高于其他气象灾害造成的损失(张强等,2014;Wilhite,2000),受到高度关注(王春乙,2007;吕娟等,2011;Turner et al., 2011;Gupta et al.,2020). ...
Plant-microbe interactions before drought influence plant physiological responses to subsequent severe drought
3
2019
... 农业干旱是农作物生长发育过程中,因降雨不足或长期无雨造成大气干旱、土壤缺水,致使作物得不到适时适量的水分补给,无法满足正常需水,影响其光合过程及生物量积累,最终造成农作物减产甚至绝收.农业干旱是影响农业生产的重要因素,也是农业生产面临的最大风险之一(王春乙,2007;肖国举和李裕,2012;张强等,2017;Mirzabaev et al.,2023).农业生产决定着粮食产量的稳定,是国家战略性、基础性核心产业,农作物种植是农业的“芯片”,是促进农业长期稳定发展,保障国家粮食安全的根本.干旱缺水直接影响农作物的生长发育和产量形成,造成粮食产量下降、品质降低,从而增加粮食生产供给的不稳定性和风险,直接威胁粮食安全,成为制约社会经济发展的重要因素之一(姚玉璧等,2013;张强等,2008;张强等,2020;IPCC,2007;IPCC,2022;Mirzabaev et al.,2023),长期以来农业旱灾对粮食安全的影响是学者和政府部门关注的重点和焦点.农业干旱对作物的影响从微观到宏观表现在细胞、生理和形态等不同层次水平上,其影响程度取决于干旱开始时间、发展和持续时间、严重程度及农作物种类和品种、所处生育期等因素(赵鸿等,2016;Ulrich et al.,2019;Gupta et al.,2020). ...
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
1
2022
... 受全球气候变化和经济快速发展的影响,全球干旱呈现多发、加重趋势,中亚地区65%的区域表现为干旱程度加剧、强度持续增强,2022年全球极端干旱问题尤为突出(李稚等,2022;张强,2022;Metz et al.,2007;Li et al.,2015;IPCC,2022).随着全球气候变暖,水循环加快,陆地蒸散发和储水量减少,干旱半干旱区不断扩张,干旱持续加剧,干旱风险进一步加大,且存在较大的区域差异(张强等,2017;Salinger et al.,2005;Cook et al.,2007;Metz et al.,2007;Huang et al.,2016).联合国《2022年全球干旱数字》报告指出,自2000年以来全球干旱次数增加了29%,全球处于高温和干旱管理的“十字路口”(UNCCD, 2022).预计到2100年,极端干旱地区的干旱次数和强度将增加1%~30%(IPCC,2007). ...
Physiological and molecular adaptations to drought in Andean potato genotypes
1
2008
... 植物生长调节剂(外用)和植物激素(内用)是影响植物生理过程的物质,其浓度很低时起调节作用,如脱落酸、乙烯、生长素、赤霉素、细胞分裂素等(Morgan,1991;康书江等,1997).在分子和生化水平上,植物通过在转录和蛋白表达过程中改变调节反应来感知和响应干旱胁迫,从而影响生化途径和代谢过程,进而影响生理和发育过程(Mane et al.,2008;Vasquez-Robinet et al.,2008;Zinselmeier et al.,1995).在土壤干旱条件下植物通常会产生脱落酸(Abscisic Acid,ABA),它是一种主要的化学根-芽胁迫信号(Davies and Zhang,1991),是通过激活对干旱响应的基因表达而表征细胞反应的一个关键信号(Chaves et al.,2003).ABA含量可作为植物抗旱性的评价指标之一,其与籽粒的生长速率有关.当土壤干旱时失水的根系产生根源信号ABA,通过木质部输送到地上部调节气孔开闭(Davies and Zhang,1991),作物根冠、叶片、花、籽粒生长进程中ABA含量明显上升(赵鸿等,2008;张玉书等,2012;Obidiegwu et al,2015),且随着干旱胁迫程度的加剧而增加,如受到水分胁迫时小麦籽粒生长减慢、结实率降低,玉米胚乳细胞分化率降低.乙烯(Ethylene)也是干旱胁迫下植物体产生的激素,在干旱感知时乙烯通过蒸腾流将其他激素信号与ABA一起发送到嫩芽中.干旱引发的乙烯释放过量,会引起小麦、玉米等籽粒产量下降(赵鸿等,2008;Obidiegwu et al.,2015). ...
2
2000
... 干旱灾害(简称“旱灾”),是指一段时间内降水量比常年明显偏少,造成某一地区经济社会活动(尤其是农业生产)和人类生活遭受较大危害的现象,普遍存在于世界各地,频繁发生于不同时期(Wilhite,2000;Li et al.,2015).旱灾是气象干旱、农业干旱、水文干旱、社会经济干旱和生态干旱之间的一种链状传递和转化过程,一个完整的干旱事件包括干旱历时、严重程度和平均强度(姚玉璧等,2013;张强等,2014;Heffernan,2013).全球每年因干旱导致的经济损失达60~80亿美元,2021年甚至高达121亿美元(American Meteorological Society, 1997;应急管理部-教育部减灾与应急管理研究院等,2022),远高于其他气象灾害造成的损失(张强等,2014;Wilhite,2000),受到高度关注(王春乙,2007;吕娟等,2011;Turner et al., 2011;Gupta et al.,2020). ...
... ;Wilhite,2000),受到高度关注(王春乙,2007;吕娟等,2011;Turner et al., 2011;Gupta et al.,2020). ...
Understanding the complex impacts of drought: a key to enhancing drought mitigation and preparedness
1
2007
... 美国、澳大利亚等受干旱影响较重的国家,在过去30 a间,率先摒弃了被动应急抗旱的方式,积极采取主动的干旱风险管理模式,即预警防旱早于应急救援(Collins et al.,2016),有效应对可能发生的干旱灾害(Fu et al.,2013;Hervás-Gámez and Delgado-Ramos,2020).具体包括3个方面:首先,通过制定抗旱预案制度、发布国家和区域干旱政策、提高干旱监测预警能力、加强干旱灾害风险评估等手段,增强全社会防御干旱的能力(Wilhite et al.,2007;Peters-lidard et al.,2021;Steinemann et al.,2015;Zarei et al.,2020);其次,在干旱易发区域或干旱发生后,采取强化各部门需水管理、普及淡水资源化以及提高水分利用效率措施,如种植抗旱作物、农业节水灌溉、废水再利用及微咸水灌溉等,也可采用工程抗旱、生物抗旱等措施尽可能降低旱灾造成的损失(WMO and GWP,2014);第三,对于干旱造成损失的,政府制定相应政策评估受损,给予救济金补贴,让受灾严重的群体尽快恢复生产生活(WMO and GWP,2017).此外,澳大利亚还利用地下含水层储水-回灌技术发展农业节水灌溉,即把冬天地表多余的水资源通过压力储存到地下100 m左右的含水层,遇到干旱月份再抽出来进行农田灌溉;通过地理信息系统(Geographic Information System,GIS)、遥感系统(Remote Sensing,RS)、全球定位系统(Global Position System,GPS)等3S系统和制图(Mapping)、监控(Monitoring)、管理(Management)系统等3M系统进行节水灌溉,以实现节约水资源的目的(https://world.qianlong.com/2022/0830/7575142.shtml). ...
National drought management policy guidelines: a template for action
1
2014
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
Benefits of action and costs of inaction: drought mitigation and preparedness
1
2017
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
Plant responses to drought and rewatering
1
2010
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
Artificial neural network modeling of leaf water potential for potatoes using RGB digital images: a greenhouse study
2
2006
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 在农业干旱监测中,农业干旱指标主要有气象指标、土壤墒情指标、作物生理生态指标和一些其他指标.气象指标包括降水距平百分数、连续无有效降水日数、湿度指标和Z指数等(孙荣强,1994;姚玉璧等,2013;国家气象中心等,2015;Zhang et al.,2009).土壤墒情指标包括土壤相对湿度、土壤有效水分存储量、土壤水分旱灾指数等(国家气象中心等,2015;Hollinger et al.,1993;Dai et al.,2004).作物生理生态指标主要是一些基于植株生长和光合特性的生理参数,用来衡量植株水分亏缺状况,如光合速率、蒸腾速率、气孔导度、复水后的光合恢复、叶水势、茎秆水势、叶绿素、叶片相对含水量、叶片扩展速率、作物水分胁迫指数、冠层温度、块茎碳同位素鉴别等(姚玉璧等,2013;张强等,2014;赵鸿等,2018;纪瑞鹏等,2019;Jefferies,1993;Medrano et al.,2002;Zakaluk and Ranjan,2006;Ricardo,2012;Ramirez et al.,2016),这些基于植株的监测指标更加方便、适时,但有些地区由于没有专用测量仪器而导致部分指标不能广泛应用. ...
Evaluation of the influence of occurrence time of drought on the annual yield of rain-fed winter wheat using backward multiple generalized estimation equation
1
2020
... 美国、澳大利亚等受干旱影响较重的国家,在过去30 a间,率先摒弃了被动应急抗旱的方式,积极采取主动的干旱风险管理模式,即预警防旱早于应急救援(Collins et al.,2016),有效应对可能发生的干旱灾害(Fu et al.,2013;Hervás-Gámez and Delgado-Ramos,2020).具体包括3个方面:首先,通过制定抗旱预案制度、发布国家和区域干旱政策、提高干旱监测预警能力、加强干旱灾害风险评估等手段,增强全社会防御干旱的能力(Wilhite et al.,2007;Peters-lidard et al.,2021;Steinemann et al.,2015;Zarei et al.,2020);其次,在干旱易发区域或干旱发生后,采取强化各部门需水管理、普及淡水资源化以及提高水分利用效率措施,如种植抗旱作物、农业节水灌溉、废水再利用及微咸水灌溉等,也可采用工程抗旱、生物抗旱等措施尽可能降低旱灾造成的损失(WMO and GWP,2014);第三,对于干旱造成损失的,政府制定相应政策评估受损,给予救济金补贴,让受灾严重的群体尽快恢复生产生活(WMO and GWP,2017).此外,澳大利亚还利用地下含水层储水-回灌技术发展农业节水灌溉,即把冬天地表多余的水资源通过压力储存到地下100 m左右的含水层,遇到干旱月份再抽出来进行农田灌溉;通过地理信息系统(Geographic Information System,GIS)、遥感系统(Remote Sensing,RS)、全球定位系统(Global Position System,GPS)等3S系统和制图(Mapping)、监控(Monitoring)、管理(Management)系统等3M系统进行节水灌溉,以实现节约水资源的目的(https://world.qianlong.com/2022/0830/7575142.shtml). ...
Comprehensive index for drought evaluation
2
2009
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 在农业干旱监测中,农业干旱指标主要有气象指标、土壤墒情指标、作物生理生态指标和一些其他指标.气象指标包括降水距平百分数、连续无有效降水日数、湿度指标和Z指数等(孙荣强,1994;姚玉璧等,2013;国家气象中心等,2015;Zhang et al.,2009).土壤墒情指标包括土壤相对湿度、土壤有效水分存储量、土壤水分旱灾指数等(国家气象中心等,2015;Hollinger et al.,1993;Dai et al.,2004).作物生理生态指标主要是一些基于植株生长和光合特性的生理参数,用来衡量植株水分亏缺状况,如光合速率、蒸腾速率、气孔导度、复水后的光合恢复、叶水势、茎秆水势、叶绿素、叶片相对含水量、叶片扩展速率、作物水分胁迫指数、冠层温度、块茎碳同位素鉴别等(姚玉璧等,2013;张强等,2014;赵鸿等,2018;纪瑞鹏等,2019;Jefferies,1993;Medrano et al.,2002;Zakaluk and Ranjan,2006;Ricardo,2012;Ramirez et al.,2016),这些基于植株的监测指标更加方便、适时,但有些地区由于没有专用测量仪器而导致部分指标不能广泛应用. ...
Ridge-furrow with full plastic film mulching improves water use efficiency and tuber yields of potato in a semiarid rainfed ecosystem
2
2014
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 第四,因地制宜推广保护性耕作措施,可同时起到保水、保土、培肥、增产的综合作用,如垄沟种植、垄沟径流集水、地膜覆盖、秸秆覆盖、免耕栽培等技术.采取多种形式的带状间作为中心的保护性耕作技术,缓解气候变暖加剧引起的水资源供求矛盾.推行麦类等条播作物与马铃薯、玉米、大豆等穴播作物或主粮作物与畜草合理间作、轮作等技术;合理套作,增加复种指数,提高耕地利用效率,如小麦套种马铃薯、小麦套种玉米等,可不同程度地提高作物产量,相当于在半干旱偏旱区的旱作农田上实现一年二熟(山仑,2011;张强等,2012a;张强等,2012b;肖国举和李裕,2012;鲍文中和周广胜,2017;Zhao et al.,2012;Zhao et al.,2014). ...
Plastic film mulch for half growing-season maximized WUE and yield of potato via moisture temperature improvement in a semi-arid agroecosystem
1
2012
... 第四,因地制宜推广保护性耕作措施,可同时起到保水、保土、培肥、增产的综合作用,如垄沟种植、垄沟径流集水、地膜覆盖、秸秆覆盖、免耕栽培等技术.采取多种形式的带状间作为中心的保护性耕作技术,缓解气候变暖加剧引起的水资源供求矛盾.推行麦类等条播作物与马铃薯、玉米、大豆等穴播作物或主粮作物与畜草合理间作、轮作等技术;合理套作,增加复种指数,提高耕地利用效率,如小麦套种马铃薯、小麦套种玉米等,可不同程度地提高作物产量,相当于在半干旱偏旱区的旱作农田上实现一年二熟(山仑,2011;张强等,2012a;张强等,2012b;肖国举和李裕,2012;鲍文中和周广胜,2017;Zhao et al.,2012;Zhao et al.,2014). ...
Global genome expression analysis of rice in response to drought and high-salinity stresses in shoot, flag leaf, and panicle
3
2007
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
... ;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
Different solute levels in two spring wheat cultivars induced by progressive field water stress at different developmental stages
4
2005
... 就干旱缺水对农业影响、干旱监测及抗旱减灾方面已进行了大量研究,从不同方面剖析了干旱胁迫后作物根、茎、叶、花、籽粒等一系列响应特征和水分传输、生理生化过程以及形态响应等(白莉萍等,2004;覃志豪等,2005;毕建杰等,2008;张强等,2015;赵鸿等,2016;McWilliam,1986;Araus et al.,2002;Chaves et al.,2003;Boyer and Westgate,2004;Zhu et al.,2005;Rampino et al.,2006;Battaglia et al.,2007;Akram,2011;Hossain et al.,2012;Boguszewska-Mankowska et al.,2018;Ulrich et al.,2019;Gupta et al.,2020),相继研发了一系列农业干旱监测指标和方法(孙荣强,1994;王春乙,2007;姚玉璧等,2013;李柏贞和周广胜,2014;刘宗元等,2014;国家气象中心等,2015;王润元等,2015;Palmer,1968;Hollinger et al.,1993;Jefferies,1993;Medrano et al.,2002;Dai et al.,2004;Zakaluk and Ranjan,2006;Zhang et al.,2009;Ricardo,2012;Ramirez et al.,2016),并从农业、水利、生物等角度制定了多种适应和减缓措施(陆亚龙和肖功建,2001;韩萍,2002;覃志豪等,2005;邓振镛等,2007;山仑,2011;肖国举和李裕,2012;熊友才和李凤民,2014;张强等,2008;张强等,2012a;张强等,2012b;周广胜等,2016;赵鸿等,2016;张强等,2017;赵鸿等,2018;Fu et al.,2013;WMO & GWP,2014;Zhao et al.,2014;WMO & GWP,2017;Qiang et al.,2019a;Gupta et al.,2020;Hervás-Gámez and Delgado-Ramos,2020).已有的大多数研究结果各有侧重,比较分散,在农业旱灾成因-影响程度-作物响应机制-监测-应对这一主线的系统性梳理和凝练方面仍需要进一步加强.鉴于农业干旱灾害的复杂性,本文在上述论述基础上,集合了农业、水利、生物、气象等各方面的研究成果,强化农业干旱灾害监测预报能力,以期为提升国家粮食安全气象服务的精细化水平提供一定的理论基础. ...
... 干旱是粮食作物生产中最主要的制约因子之一,在干旱半干旱区表现尤其显著(王春玲等,2017;Chaves et al.,2003;Boyer and Westgate,2004;Bandi et al.,2012;Hossain et al.,2012).作物在生长季一段时间或多段时间内由于供水不足往往遭受不同程度和不同持续时间的干旱(刘宗元等,2014;McWilliam,1986;Gambetta et al.,2020),无论是依照严重程度划分的轻旱、中旱、重旱、特旱,还是从持续时间上划分的间歇性(断断续续)干旱和持续性干旱,都会对作物造成不同程度的伤害和影响.作物对干旱信号感知和传导后,通过分子、细胞、生理等不同水平层次上影响作物的形态、结构和功能(Ulrich et al.,2019;Gupta et al.,2020),这涉及作物的生长状况、生理生化及代谢过程、形态建成、产量形成等各种生物过程.干旱对作物的影响程度取决于干旱开始时间、发展速度、持续时长、环境条件及作物物种、生育期(苗期、拔节期、开花期、灌浆期、成熟期等)、生长阶段(营养生长和生殖生长)等(张强等,2015;Araus et al.,2002;Hirt and Shinozaki,2004;Zhu et al.,2005;Akram,2011;Boguszewska-Mankowska et al.,2018),还受作物器官、组织、细胞、亚细胞以及基因型等影响(赵鸿等,2016;Rampino et al.,2006;Zhou et al.,2007;Battaglia et al.,2007;Pinheiro and Chaves,2011;Qin et al.,2019).图7描述了不同类型干旱对农作物的影响机理及作物在形态、生理和分子水平上对干旱缺水的响应机制. ...
... 不断变化的干旱扰乱了作物的生长周期,作物通过土壤供应水分进行光合作用制造有机物,有机物积累量的大小往往表现在植株的株高、茎粗、根冠、叶色、叶面积和产量形成等一系列动态变化过程中(雷俊等,2017;Boyer and Westgate,2004;Rampino et al.,2006;Loyla Rodríguez et al.,2016;Hirut et al.,2017).水分亏缺会减缓植物生长发育,导致器官变小,阻碍花的发育和籽粒的灌浆,根是作物适应干旱最主要的器官(Ricardo,2012).在作物形态方面,轻度干旱胁迫下作物水分供给不足,生物量向根部积聚,地下部分功能根的数量增多、长度和根重增加、块茎类作物的起芽量减少;随着干旱胁迫程度的增加,植株-根系的水分关系不断被扰乱(Farooq et al.,2009;Ricardo,2012),植株生长速率大幅下降,叶片数量减少和尺寸减小、光合叶面积减小、顶端分生组织细胞分裂减慢甚至停止、茎尖伸长区细胞伸长受到抑制、植株茎杆节间缩短、个体矮小(Zhu et al.,2005;Farooq et al.,2009;Akram,2011),同时叶片内膨压降低,叶肉细胞扩展受限,致使叶片组织疲软、卷曲、萎蔫、枯黄甚至死亡(白莉萍等,2004;Nicolas et al.,1985).如小麦、玉米等禾谷类作物在拔节期和抽穗期遇到水分供应不足时,植株节间细胞扩展受限,导致植株低矮,甚至可能不抽穗;在灌浆期遭受水分胁迫时,则引起灌浆时间缩短、灌浆速率降低、植株衰老提早,导致作物穗数、穗粒数、粒重等构成要素降低,最终造成籽粒产量和品质下降(张军等,2014;赵鸿等,2016;EI Hafid et al.,1998;Lesk et al.,2016).此外,干旱在更大程度上使植株生长受阻,水分利用效率显著降低,如马铃薯早期的干旱胁迫显著降低了水分的利用效率,大大减少了植株生长及其生物量的累积(Costa et al.,1997;Farooq et al.,2009).间歇性干旱的发生和历时对作物生物量和产量的影响可能比干旱强度的影响大得多,取决于胁迫持续时间和物候期(Serraj et al.,2004),这间接降低了光合速率、CO2固定,最终导致同化产物减少(Mafakheri et al.,2010;Pinheiro and Chaves,2011). ...
... 干旱对作物的主要影响是光合作用减少,这是由叶片扩张减少、光合机制受损、叶片过早衰老等引起的(Ambavaram et al.,2014).土壤干旱或供水不足会直接影响植株冠层、叶片及其光合过程,缺水时作物根系吸收水分首先受到限制,向地上部分运送的水分不断减少,地上冠层部分生长减缓,干物质积累量减少(Zhu et al.,2005;Akram,2011),根冠比(地下部分与地上部分的比值)增加(Zhou et al.,2007).一方面,土壤水分减少引起叶片水势下降、叶片含水量降低、保卫细胞压力势减小而散失水分,使得气孔开度减小甚至闭合,阻碍CO2气体交换,导致光合速率下降(毕建杰等,2008;Battaglia et al.,2007;Zhou et al.,2007);另一方面,干旱胁迫时叶片生长减缓,光合叶面积减小,淀粉水解作用增强,糖类积累增大,光合产物向汇器官运输受限,呼吸作用加强,呼吸消耗增大(赵鸿等,2008;赵鸿等,2016;Kobata et al.,1992),最终导致作物生产力下降(Jones and Corlett,1992).研究表明,当土壤水分在作物可利用水分的50%以上时,作物可获得最大的产量(Mackerron and Jelferies,1986).在作物生理水平上,干旱导致的作物光合作用减少是通过气孔限制和非气孔限制引起的,在轻度至中度干旱胁迫下叶片的气孔开放和闭合受到影响,气孔因素占主导作用,抑制生物量的积累;在严重干旱胁迫下非气孔因素占主导地位(Angelopoulos et al.,1996),限制了光化学和酶活性,损伤叶绿体中光抑制,从而影响生理过程和生化代谢(Baker,2008;Xu et al.,2010).此外,在干旱胁迫下气孔关闭会降低叶肉中CO2可用性,电子传递和生化途径变化会导致光合作用减少(Boyer,1976;Cornic et al.,1983;Genty et al.,1987;Obidiegwu et al.,2015). ...
Low water potential disrupts carbohydrate metabolism in maize (Zea mays L.) ovaries
1
1995
... 植物生长调节剂(外用)和植物激素(内用)是影响植物生理过程的物质,其浓度很低时起调节作用,如脱落酸、乙烯、生长素、赤霉素、细胞分裂素等(Morgan,1991;康书江等,1997).在分子和生化水平上,植物通过在转录和蛋白表达过程中改变调节反应来感知和响应干旱胁迫,从而影响生化途径和代谢过程,进而影响生理和发育过程(Mane et al.,2008;Vasquez-Robinet et al.,2008;Zinselmeier et al.,1995).在土壤干旱条件下植物通常会产生脱落酸(Abscisic Acid,ABA),它是一种主要的化学根-芽胁迫信号(Davies and Zhang,1991),是通过激活对干旱响应的基因表达而表征细胞反应的一个关键信号(Chaves et al.,2003).ABA含量可作为植物抗旱性的评价指标之一,其与籽粒的生长速率有关.当土壤干旱时失水的根系产生根源信号ABA,通过木质部输送到地上部调节气孔开闭(Davies and Zhang,1991),作物根冠、叶片、花、籽粒生长进程中ABA含量明显上升(赵鸿等,2008;张玉书等,2012;Obidiegwu et al,2015),且随着干旱胁迫程度的加剧而增加,如受到水分胁迫时小麦籽粒生长减慢、结实率降低,玉米胚乳细胞分化率降低.乙烯(Ethylene)也是干旱胁迫下植物体产生的激素,在干旱感知时乙烯通过蒸腾流将其他激素信号与ABA一起发送到嫩芽中.干旱引发的乙烯释放过量,会引起小麦、玉米等籽粒产量下降(赵鸿等,2008;Obidiegwu et al.,2015). ...


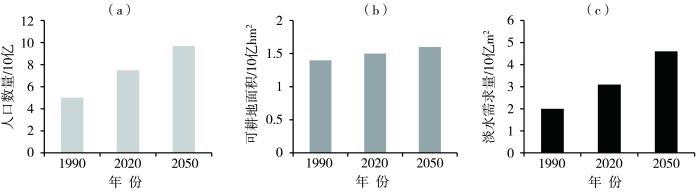
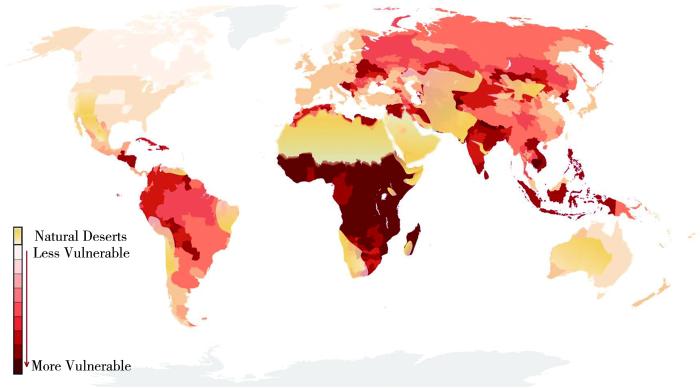
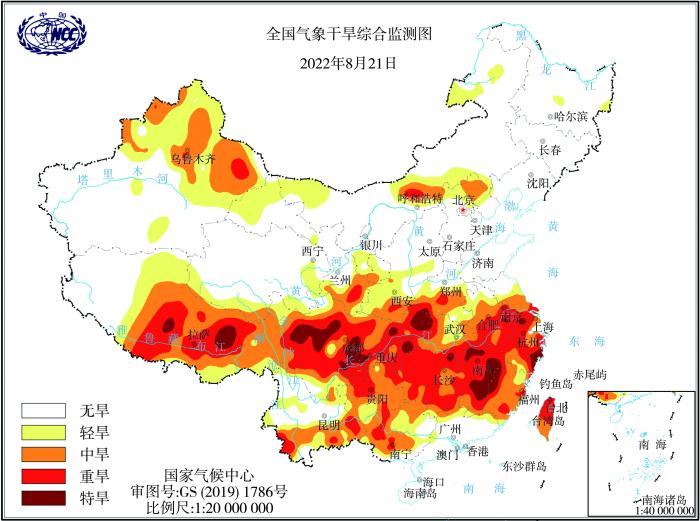
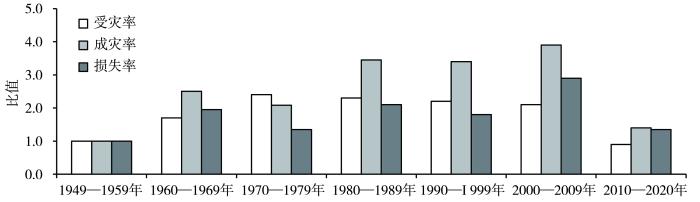
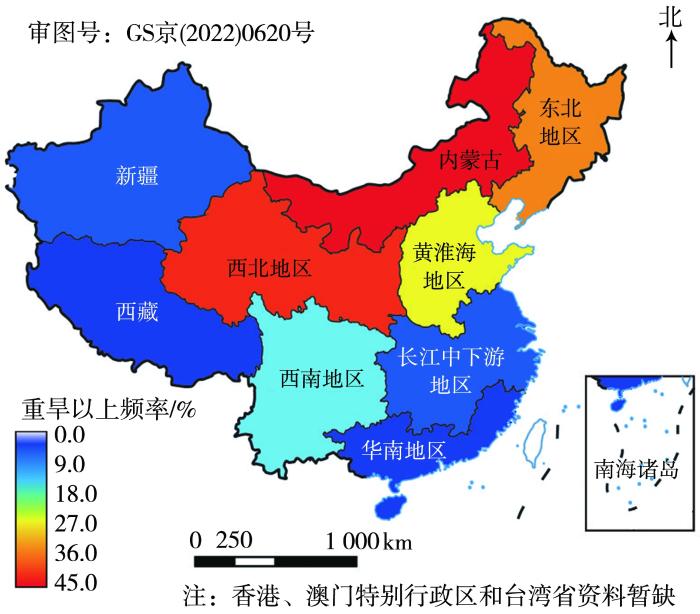

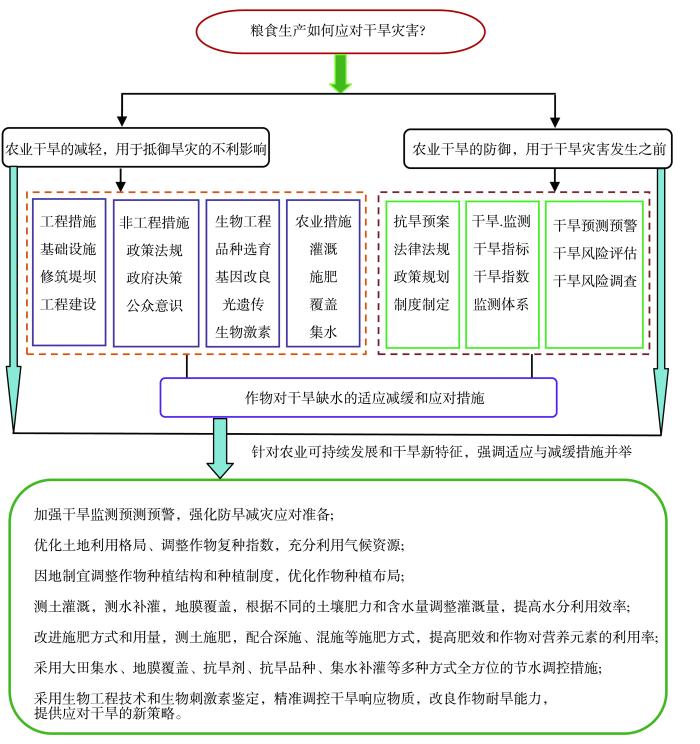



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}