Photosynthetic characteristics and response mechanism of spring maize at seven-leaf stage under drought stress
TANG Yurui,1,2,3,4, QI Yue,1,2,3,4, WANG Heling1,2,3,4, YANG Yang1,2,3,4, ZHAO Hong1,2,3,4, ZHANG Kai1,2,3,4, WEI Xingxing1,2,3,4, WANG Renkui5
1. Institute of Arid Meteorology, China Meteorological Administration, Key Laboratory of Arid Climatic Change and Reducing Disaster of Gansu Province, Key Laboratory of Arid Climatic Change and Disaster Reduction of CMA, Lanzhou 730020, China
2. Dingxi Drought Meteorology and Ecological Environment Field Scientific Experimental Station of CMA, Dingxi 743000, Gansu, China
3. Wuwei National Climate Observatory of China Meteorological Administration, Wuwei 733000, Gansu, China
4. Zhangye National Climate Observatory of China Meteorological Administration, Zhangye 734000, Gansu, China
5. Jingyuan Meteorological Bureau of Gansu Province, Jingyuan 730600, Gansu, China
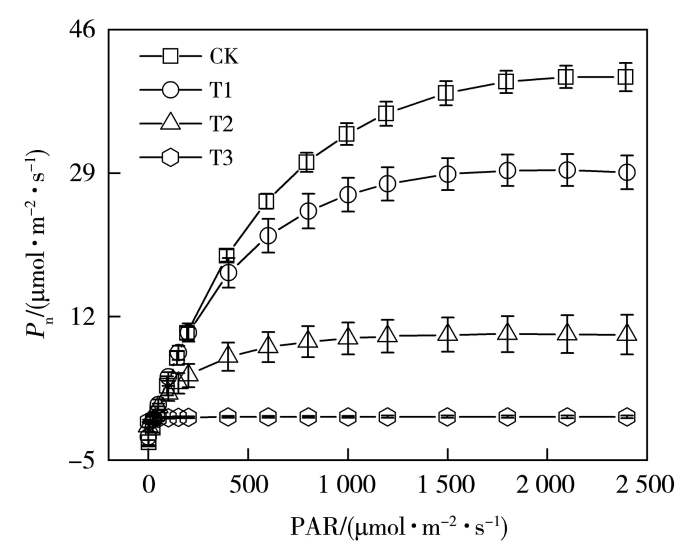
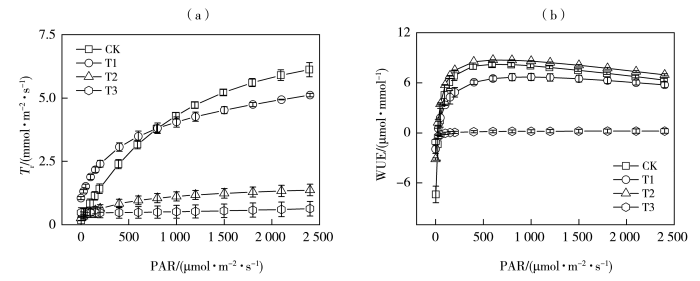
干旱胁迫是限制作物生长的主要因素,研究春玉米(Zea mays L.)苗期光合特性与生理抗旱机制,对有效抗御干旱、促进作物产量形成等方面具有重要意义。以春玉米为研究对象,采用盆栽试验,设置对照处理(简称“CK”处理)、轻度干旱(T1)、中度干旱(T2)、重度干旱(T3)4种梯度,研究不同干旱程度下春玉米七叶期叶片的光合生理特征及其抗旱机制。结果表明:干旱显著降低了春玉米叶片的净光合速率(Pn)及最大净光合速率(Pnmax);光响应曲线的初始斜率(α0)、暗呼吸速率(Rd)、表观量子效率(Apparent Quantum Efficiency,AQE)在中度、重度干旱处理下显著下降,光饱和系数(γ0)显著增加,叶片的光能利用率显著下降;蒸腾速率(Tr)随着干旱胁迫的加剧逐渐降低,水分利用效率(Water Use Efficiency,WUE)在中度干旱处理下显著增加,重度干旱下显著降低;气孔导度(Gs)随着干旱胁迫的加剧逐渐下降,轻度、中度干旱处理下净光合速率下降的因素以气孔限制为主,轻度干旱下存在气孔非均匀关闭现象,重度干旱下胞间CO2浓度(Ci)显著上升,气孔限制值(Ls)显著降低,以非气孔限制为主导因素。
关键词:干旱胁迫;
气体交换参数;
水分利用效率;
响应
Abstract
Drought stress is a major limiting factor for crop growth. Investigating the photosynthetic characteristics and physiological drought resistance mechanisms of spring maize (Zea mays L.) at the seedling stage is crucial for enhancing drought resilience and promoting crop yield. In this study, taking spring maize as the research object and using pot experiments, four gradients of control treatment (referred to as “CK” treatment), mild drought (T1), moderate drought (T2), and severe drought (T3) were set up to study the photosynthetic physiological characteristics and drought resistance mechanism at the seven-leaf stage of spring maize under different drought degrees. The results show that drought stress significantly reduced both the net photosynthetic rate (Pn) and the maximum net photosynthetic rate (Pnmax) of spring maize leaves. Under moderate and severe drought conditions, the initial slope (α0), the dark respiration rate (Rd), and the apparent quantum efficiency (AQE) of the light response curve decreased significantly, while the light saturation coefficient (γ0) increased markedly, indicating the light energy utilization rate of the leaves decreased significantly. The transpiration rate (Tr) gradually decreased with increasing of drought intensity, the water use efficiency (WUE) significantly enhanced under moderate drought but sharply reduced under severe drought. Stomatal conductance (Gs) progressively declined with drought stress intensification. Stomatal limitation was identified as the primary factor reducing photosynthetic rate under mild and moderate drought conditions, with non-uniform stomatal closure phenomenon under mild drought. Under severe drought condition, intercellular CO2 concentration (Ci) significantly increased, and the stomatal limitation value (Ls) decreased dramatically, demonstrating the dominance of non-stomatal limitation.
Keywords:drought stress;
gas exchange parameters;
water use efficiency;
response
TANG Yurui, QI Yue, WANG Heling, YANG Yang, ZHAO Hong, ZHANG Kai, WEI Xingxing, WANG Renkui. Photosynthetic characteristics and response mechanism of spring maize at seven-leaf stage under drought stress[J]. Arid Meteorology, 2025, 43(2): 176-185 DOI:10.11755/j.issn.1006-7639-2025-02-0176
0 引言
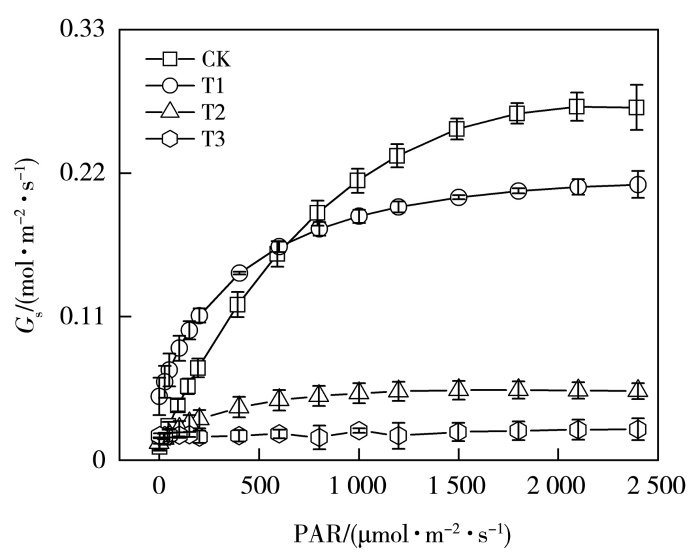
光合作用是地球生态系统一切生命活动的能量来源,也是作物产量形成的生理基础,其对环境变化十分敏感(卜令铎等,2010;李建查等,2018)。作为植物最常见的生长逆境,干旱是抑制玉米生长和光合作用下降的重要原因(张仁和等,2011),限制着植物的光能利用率,甚至造成叶片结构的损伤(Mukarram et al.,2021),最终导致植物生物量及产量的下降(Jia et al.,2020)。叶片光合作用对光的响应曲线是研究作物光合作用对环境变化响应的重要手段(李义博等,2017),可反映植物对不同光强的利用规律,并指示植物的生长情况及其抗逆性。通常以气孔导度、净光合速率、蒸腾速率及水分利用效率等指标来表征作物的光合特性(于文颖等,2015)。气孔不仅是水分和光合原料进入细胞的关键通道,在干旱感知和信号传导过程中也发挥着不可或缺的作用(Laxa et al.,2019)。Hetherington和Woodward(2003)研究指出,气孔面积仅占叶片面积的5%,但经由气孔蒸发的水分却高达70%。由此可见,气孔调节作为植物抵御胁迫环境的核心机制,对于维持植物体内水分平衡、保障光合作用正常进行意义重大。气孔导度作为衡量气孔变化特征的重要指标,其大小直接影响叶片中CO2的交换速率和水分的蒸腾速率(龚容和高琼,2015)。适度水分亏缺下叶片可通过改变气孔的形态和气孔导度的大小来控制叶片内部CO2浓度和蒸腾速率,协调碳同化和水分消耗之间的关系(Ennahli and Earl,2005),同时影响叶片水分利用效率(Water Use Efficiency,WUE),最终形成高效的抗旱策略以适应干旱环境(李民青等,2023),随着干旱条件的进一步发展,作物生产可能出现环境约束向生理生态约束的转变,植物气孔可能关闭,导致叶绿体结构的变化和对植物膜系统的损害,因而阐明光合速率下降的生理机制对于准确识别作物干旱程度和制定对策至关重要(Song et al.,2020)。
植物光合作用主要受气孔和非气孔因素影响(Jia et al.,2020),前者指干旱胁迫导致的气孔导度降低,CO2进入叶片的通道受阻而使光合速率下降(Song et al.,2020),后者主要是叶肉细胞电子传递和光合磷酸化受阻,暗反应中相关酶活性降低,导致光合速率下降(Farquhar and Sharkey,1982)。宋丰萍和蒙祖庆(2018)研究表明,随着干旱胁迫的逐步加剧、持续时间不断延长,气孔和非气孔因素往往会同时对植物光合作用产生影响,在这一过程中,非气孔因素最终会成为主导限制因子。基于此,麻雪艳和周广胜(2018)提出,气孔限制向非气孔限制的转折点,可作为判定玉米是否受到严重干旱胁迫的重要依据。
干旱是农作物生长的主要制约因素之一(Lobell et al.,2014),玉米作为中国种植范围最广的谷类作物,在农业及国民经济中占据着重要地位(宋贺等,2019)。全球气候变化背景下水热格局的改变,加剧了干旱等非生物胁迫因子对作物生长和产量的限制作用(李建查等,2018)。近年来,光照、水分胁迫等环境因子对作物光合生理特征的影响得到了广泛关注(张仁和等,2011;Sharma and Zheng,2019;姜鹏等,2023),不同强度干旱过程对玉米生长发育、光合生理的影响也有较多研究,主要集中在拔节期和抽雄期(于文颖等,2015;宋贺等,2019;杨阳等,2022)。萌发期和苗期(前期干旱)是春玉米干旱发生频率最高的阶段(任宗悦等,2020),特别是玉米营养生长阶段的七叶期(齐月等,2023),此时遭遇干旱会严重抑制春玉米的生长发育(卜令铎等,2010;Song et al.,2019)。然而,目前针对干旱胁迫下春玉米七叶期的光合响应特征及其影响机制的研究尚显不足,因此本文通过研究春玉米七叶期光合参数变化特征及其对干旱的响应,试图明晰不同干旱程度下春玉米七叶期的叶片生理抗旱机制,以期为农业生产过程中春玉米生长前期的水分管理提供理论依据。
Fig.2
The light-response curves of transpiration rate (a) and instantaneous water use efficiency (b) of spring maize leaves under CK treatment and different drought stress conditions
Tab.2
表2
表2CK处理和不同干旱胁迫下春玉米叶片瞬时水分利用效率对光的响应参数
Tab.2 The light-response parameters of the instantaneous water use efficiency of spring maize leaves under CK treatment and different drought stress conditions
Fig.3
The variation characteristics of leaf water potential of spring maize under CK treatment and different drought stress conditions at 06:00 (a) and 10:00 (b)
Fig.5
The light-response curves of intercellular CO2 concentration (a) and stomatal limit value (b) of spring maize leaves under CK treatment and different drought stress conditions
Tab.4
表4
表4CK处理和不同干旱胁迫下春玉米气孔限制值对光的响应参数
Tab.4 The light-response parameters of stomatal limit value of spring maize leaves under CK treatment and different drought stress conditions
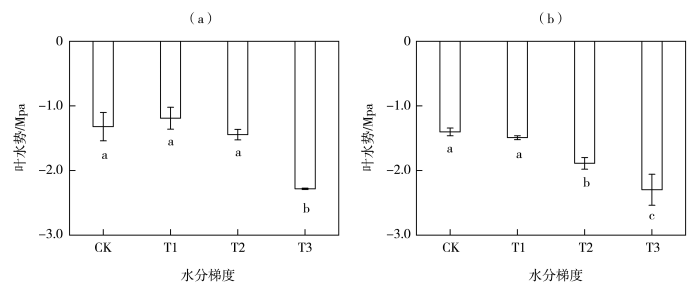
水分利用效率(WUE)是植物基本的生理生态特征,高效的WUE有利于维持植物的碳水平衡和适应干旱环境(李民青等,2023)。叶片尺度的WUE由光合作用与蒸腾作用共同决定,因而影响作物这两个生理过程的因素均会引起叶片WUE的变化(姜寒冰等,2019)。本研究中,随着干旱胁迫的增加,玉米叶片的Pn和Tr均逐步降低,WUE最终表现为中旱与对照无显著差异,轻旱和重旱显著降低,10:00测量的叶水势,在轻旱与对照下无显著差异,中、重旱下显著降低。叶水势能够直接反映作物的水分亏缺情况,叶水势对WUE的影响是通过影响气孔行为实现的,当干旱胁迫刚开始时叶片可通过气孔的部分关闭来减少水分丧失,从而使叶水势在一定范围内保持恒定(张文丽等,2006),因而叶水势在轻旱处理下与对照处理无明显差异。但叶水势发生轻微下降都将提高植物激素脱落酸(Abscisic Acid,ABA)浓度,引起叶片气孔的轻微关闭以防止蒸腾性水分流失(Laxa et al.,2019),即气孔导度降低在叶片水势还没有明显变化之前就已经发生,这种生存策略会减少叶片CO2的吸收量,最终导致净光合速率下降,从而轻旱处理下WUE小于对照处理。当植物遭受较强的干旱胁迫后,会优先考虑生存而非生产力,而角质层损失的水量决定其存活能力(Mukarram et al.,2021),气孔的关闭不仅为了防止植物失水过度,同时可以使叶片水分利用效率最大化(战吉宬等,2005)。叶水势在中旱下显著减小,气孔进一步关闭,以水分损失最小化为主要目的使蒸腾速率急剧降低(赵福年等,2019),此时蒸腾对干旱的响应比光合反应更强烈,因此蒸腾速率下降幅度要快于光合速率,也正是光合作用和蒸腾速率间的耦合关系促使中旱胁迫下水分利用效率最高,即气孔导度对叶片在碳同化和水分耗散的调节中符合最优控制的原则,这是植物抗旱策略的重要组成部分。重旱胁迫会导致叶绿素功能障碍并破坏光合器官(Cai et al.,2020),气孔对叶水势失去调节能力,因而重旱下叶水势降到极低,WUE显著减小,这与蔡福等(2017)的研究一致。
3.3 干旱胁迫下春玉米光合作用的影响因素
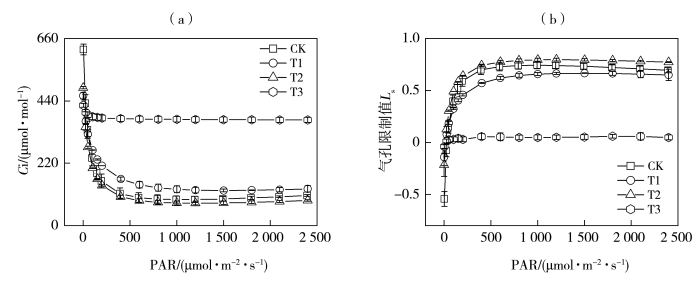
植物受到干旱胁迫时光合作用被限制,碳同化能力下降(Neto et al.,2017),光合速率的下降由气孔和非气孔因素共同决定(Jia et al.,2020)。叶子飘等(2019)研究表明,当所有气孔开闭行为保持一致时,判断叶片光合速率下降是否由气孔因素主导,需依据胞间CO2浓度(Ci)和气孔限制值的变化趋势来判定,而非单纯关注Ci的下降幅度。根据Farquhar和Sharkey(1982)的观点,在Pn减小的情况下,Ci下降且Ls升高时,气孔限制占主导作用,当Ci升高且Ls下降时,非气孔因素占主导作用。本研究中,随着光照增强,轻旱胁迫下玉米Gs相较于对照处理显著下降,叶片Ci增加,Ls减小,根据Ci的变化方向判断此时以非气孔限制为主导因素使净光合速率降低。但许大全(2013)指出叶片气孔的不均关闭现象会使Ci恒定不变,水分胁迫条件下容易造成非气孔限制的假象,当作物突然遭受水分胁迫,植物为维持正常的叶水势,通过关闭部分气孔的方式减少水分蒸腾(Sharma and Zheng,2019),计算的Ci会高于或者等于实际值,进而造成非气孔限制的假象。Medrano等(2002)提出,可由光饱和的气孔导度作为干旱条件下光合作用的气孔和非气孔限制之间的拐点指标,最大气孔导度值在0.05 mol·m-2·s-1左右时即为拐点值,因而轻旱下主要是气孔限制影响了叶片碳同化能力,导致光合速率下降。中旱处理下玉米叶片Ci减小,Ls增加,Gs和Pn均显著下降,结合最大气孔导度拐点值,此时叶片净光合速率下降的因素以气孔限制为主导,非气孔限制逐渐增强。中旱处理下植物气孔的开放程度进一步降低,气孔阻力增加,限制了光合作用原料的供应,导致Ci降低(Song et al.,2020),最终使作物光合作用速率降低,叶片通过气孔调节的方式降低水分蒸腾,避免干旱对叶片结构的损伤,这可能是本文中旱处理下气孔限制始终占主导因素的原因,体现了植物适应胁迫环境的气孔调节机制,表明玉米属较耐旱的作物,在受到干旱胁迫后作物光合系统的自身调节能力较强,可忍耐一定程度的水分亏缺(陈图强等,2023)。发生轻微干旱时,植物的抗氧化防御机制一般可以及时清除活性氧自由基以避免膜损伤(Jia et al.,2020),但重旱下土壤水分胁迫超出了植物可应对的阈值,叶片不能得到生存所需的最低水分,植物收支平衡被打破,叶肉细胞光合活性降低,抗氧化防御机理极其脆弱,活性氧的积累会破坏光系统(Sharma and Zheng,2019),植物无法维持正常生理活动。重旱下玉米叶片Ci显著增加,Ls明显减小,非气孔因素限制作物的光合作用,因而净光合速率等光合参数趋近于零。
A key question for climate change adaptation is whether existing cropping systems can become less sensitive to climate variations. We use a field-level data set on maize and soybean yields in the central United States for 1995 through 2012 to examine changes in drought sensitivity. Although yields have increased in absolute value under all levels of stress for both crops, the sensitivity of maize yields to drought stress associated with high vapor pressure deficits has increased. The greater sensitivity has occurred despite cultivar improvements and increased carbon dioxide and reflects the agronomic trend toward higher sowing densities. The results suggest that agronomic changes tend to translate improved drought tolerance of plants to higher average yields but not to decreasing drought sensitivity of yields at the field scale.
MEDRANOH, ESCALONAJ M, BOTAJ, et al, 2002.
Regulation of photosynthesis of C3 plants in response to progressive drought: Stomatal conductance as a reference parameter
NETOM C L, CERQUEIRAJ V A, DACUNHA J R, et al, 2017.
Cyclic electron flow, NPQ and photorespiration are crucial for the establishment of young plants of Ricinus communis and Jatropha curcas exposed to drought
Quantifying light response of leaf-scale water use efficiency and its interrelationships with photosynthesis and stomatal conductance in C3 and C4 species
[J]. Frontiers in Plant Science, 11: 374. DOI: 10.3389/fpls.2020.00374.
... 光合作用是地球生态系统一切生命活动的能量来源,也是作物产量形成的生理基础,其对环境变化十分敏感(卜令铎等,2010;李建查等,2018).作为植物最常见的生长逆境,干旱是抑制玉米生长和光合作用下降的重要原因(张仁和等,2011),限制着植物的光能利用率,甚至造成叶片结构的损伤(Mukarram et al.,2021),最终导致植物生物量及产量的下降(Jia et al.,2020).叶片光合作用对光的响应曲线是研究作物光合作用对环境变化响应的重要手段(李义博等,2017),可反映植物对不同光强的利用规律,并指示植物的生长情况及其抗逆性.通常以气孔导度、净光合速率、蒸腾速率及水分利用效率等指标来表征作物的光合特性(于文颖等,2015).气孔不仅是水分和光合原料进入细胞的关键通道,在干旱感知和信号传导过程中也发挥着不可或缺的作用(Laxa et al.,2019).Hetherington和Woodward(2003)研究指出,气孔面积仅占叶片面积的5%,但经由气孔蒸发的水分却高达70%.由此可见,气孔调节作为植物抵御胁迫环境的核心机制,对于维持植物体内水分平衡、保障光合作用正常进行意义重大.气孔导度作为衡量气孔变化特征的重要指标,其大小直接影响叶片中CO2的交换速率和水分的蒸腾速率(龚容和高琼,2015).适度水分亏缺下叶片可通过改变气孔的形态和气孔导度的大小来控制叶片内部CO2浓度和蒸腾速率,协调碳同化和水分消耗之间的关系(Ennahli and Earl,2005),同时影响叶片水分利用效率(Water Use Efficiency,WUE),最终形成高效的抗旱策略以适应干旱环境(李民青等,2023),随着干旱条件的进一步发展,作物生产可能出现环境约束向生理生态约束的转变,植物气孔可能关闭,导致叶绿体结构的变化和对植物膜系统的损害,因而阐明光合速率下降的生理机制对于准确识别作物干旱程度和制定对策至关重要(Song et al.,2020). ...
... 干旱是农作物生长的主要制约因素之一(Lobell et al.,2014),玉米作为中国种植范围最广的谷类作物,在农业及国民经济中占据着重要地位(宋贺等,2019).全球气候变化背景下水热格局的改变,加剧了干旱等非生物胁迫因子对作物生长和产量的限制作用(李建查等,2018).近年来,光照、水分胁迫等环境因子对作物光合生理特征的影响得到了广泛关注(张仁和等,2011;Sharma and Zheng,2019;姜鹏等,2023),不同强度干旱过程对玉米生长发育、光合生理的影响也有较多研究,主要集中在拔节期和抽雄期(于文颖等,2015;宋贺等,2019;杨阳等,2022).萌发期和苗期(前期干旱)是春玉米干旱发生频率最高的阶段(任宗悦等,2020),特别是玉米营养生长阶段的七叶期(齐月等,2023),此时遭遇干旱会严重抑制春玉米的生长发育(卜令铎等,2010;Song et al.,2019).然而,目前针对干旱胁迫下春玉米七叶期的光合响应特征及其影响机制的研究尚显不足,因此本文通过研究春玉米七叶期光合参数变化特征及其对干旱的响应,试图明晰不同干旱程度下春玉米七叶期的叶片生理抗旱机制,以期为农业生产过程中春玉米生长前期的水分管理提供理论依据. ...
关键发育期干旱及复水过程对春玉米主要生理参数的影响
1
2017
... 水分利用效率(WUE)是植物基本的生理生态特征,高效的WUE有利于维持植物的碳水平衡和适应干旱环境(李民青等,2023).叶片尺度的WUE由光合作用与蒸腾作用共同决定,因而影响作物这两个生理过程的因素均会引起叶片WUE的变化(姜寒冰等,2019).本研究中,随着干旱胁迫的增加,玉米叶片的Pn和Tr均逐步降低,WUE最终表现为中旱与对照无显著差异,轻旱和重旱显著降低,10:00测量的叶水势,在轻旱与对照下无显著差异,中、重旱下显著降低.叶水势能够直接反映作物的水分亏缺情况,叶水势对WUE的影响是通过影响气孔行为实现的,当干旱胁迫刚开始时叶片可通过气孔的部分关闭来减少水分丧失,从而使叶水势在一定范围内保持恒定(张文丽等,2006),因而叶水势在轻旱处理下与对照处理无明显差异.但叶水势发生轻微下降都将提高植物激素脱落酸(Abscisic Acid,ABA)浓度,引起叶片气孔的轻微关闭以防止蒸腾性水分流失(Laxa et al.,2019),即气孔导度降低在叶片水势还没有明显变化之前就已经发生,这种生存策略会减少叶片CO2的吸收量,最终导致净光合速率下降,从而轻旱处理下WUE小于对照处理.当植物遭受较强的干旱胁迫后,会优先考虑生存而非生产力,而角质层损失的水量决定其存活能力(Mukarram et al.,2021),气孔的关闭不仅为了防止植物失水过度,同时可以使叶片水分利用效率最大化(战吉宬等,2005).叶水势在中旱下显著减小,气孔进一步关闭,以水分损失最小化为主要目的使蒸腾速率急剧降低(赵福年等,2019),此时蒸腾对干旱的响应比光合反应更强烈,因此蒸腾速率下降幅度要快于光合速率,也正是光合作用和蒸腾速率间的耦合关系促使中旱胁迫下水分利用效率最高,即气孔导度对叶片在碳同化和水分耗散的调节中符合最优控制的原则,这是植物抗旱策略的重要组成部分.重旱胁迫会导致叶绿素功能障碍并破坏光合器官(Cai et al.,2020),气孔对叶水势失去调节能力,因而重旱下叶水势降到极低,WUE显著减小,这与蔡福等(2017)的研究一致. ...
不同灌水量对核桃树生理、生长和果实品质的影响
1
2023
... 植物受到干旱胁迫时光合作用被限制,碳同化能力下降(Neto et al.,2017),光合速率的下降由气孔和非气孔因素共同决定(Jia et al.,2020).叶子飘等(2019)研究表明,当所有气孔开闭行为保持一致时,判断叶片光合速率下降是否由气孔因素主导,需依据胞间CO2浓度(Ci)和气孔限制值的变化趋势来判定,而非单纯关注Ci的下降幅度.根据Farquhar和Sharkey(1982)的观点,在Pn减小的情况下,Ci下降且Ls升高时,气孔限制占主导作用,当Ci升高且Ls下降时,非气孔因素占主导作用.本研究中,随着光照增强,轻旱胁迫下玉米Gs相较于对照处理显著下降,叶片Ci增加,Ls减小,根据Ci的变化方向判断此时以非气孔限制为主导因素使净光合速率降低.但许大全(2013)指出叶片气孔的不均关闭现象会使Ci恒定不变,水分胁迫条件下容易造成非气孔限制的假象,当作物突然遭受水分胁迫,植物为维持正常的叶水势,通过关闭部分气孔的方式减少水分蒸腾(Sharma and Zheng,2019),计算的Ci会高于或者等于实际值,进而造成非气孔限制的假象.Medrano等(2002)提出,可由光饱和的气孔导度作为干旱条件下光合作用的气孔和非气孔限制之间的拐点指标,最大气孔导度值在0.05 mol·m-2·s-1左右时即为拐点值,因而轻旱下主要是气孔限制影响了叶片碳同化能力,导致光合速率下降.中旱处理下玉米叶片Ci减小,Ls增加,Gs和Pn均显著下降,结合最大气孔导度拐点值,此时叶片净光合速率下降的因素以气孔限制为主导,非气孔限制逐渐增强.中旱处理下植物气孔的开放程度进一步降低,气孔阻力增加,限制了光合作用原料的供应,导致Ci降低(Song et al.,2020),最终使作物光合作用速率降低,叶片通过气孔调节的方式降低水分蒸腾,避免干旱对叶片结构的损伤,这可能是本文中旱处理下气孔限制始终占主导因素的原因,体现了植物适应胁迫环境的气孔调节机制,表明玉米属较耐旱的作物,在受到干旱胁迫后作物光合系统的自身调节能力较强,可忍耐一定程度的水分亏缺(陈图强等,2023).发生轻微干旱时,植物的抗氧化防御机制一般可以及时清除活性氧自由基以避免膜损伤(Jia et al.,2020),但重旱下土壤水分胁迫超出了植物可应对的阈值,叶片不能得到生存所需的最低水分,植物收支平衡被打破,叶肉细胞光合活性降低,抗氧化防御机理极其脆弱,活性氧的积累会破坏光系统(Sharma and Zheng,2019),植物无法维持正常生理活动.重旱下玉米叶片Ci显著增加,Ls明显减小,非气孔因素限制作物的光合作用,因而净光合速率等光合参数趋近于零. ...
叶片结构的水力学特性对植物生理功能影响的研究进展
1
2015
... 光合作用是地球生态系统一切生命活动的能量来源,也是作物产量形成的生理基础,其对环境变化十分敏感(卜令铎等,2010;李建查等,2018).作为植物最常见的生长逆境,干旱是抑制玉米生长和光合作用下降的重要原因(张仁和等,2011),限制着植物的光能利用率,甚至造成叶片结构的损伤(Mukarram et al.,2021),最终导致植物生物量及产量的下降(Jia et al.,2020).叶片光合作用对光的响应曲线是研究作物光合作用对环境变化响应的重要手段(李义博等,2017),可反映植物对不同光强的利用规律,并指示植物的生长情况及其抗逆性.通常以气孔导度、净光合速率、蒸腾速率及水分利用效率等指标来表征作物的光合特性(于文颖等,2015).气孔不仅是水分和光合原料进入细胞的关键通道,在干旱感知和信号传导过程中也发挥着不可或缺的作用(Laxa et al.,2019).Hetherington和Woodward(2003)研究指出,气孔面积仅占叶片面积的5%,但经由气孔蒸发的水分却高达70%.由此可见,气孔调节作为植物抵御胁迫环境的核心机制,对于维持植物体内水分平衡、保障光合作用正常进行意义重大.气孔导度作为衡量气孔变化特征的重要指标,其大小直接影响叶片中CO2的交换速率和水分的蒸腾速率(龚容和高琼,2015).适度水分亏缺下叶片可通过改变气孔的形态和气孔导度的大小来控制叶片内部CO2浓度和蒸腾速率,协调碳同化和水分消耗之间的关系(Ennahli and Earl,2005),同时影响叶片水分利用效率(Water Use Efficiency,WUE),最终形成高效的抗旱策略以适应干旱环境(李民青等,2023),随着干旱条件的进一步发展,作物生产可能出现环境约束向生理生态约束的转变,植物气孔可能关闭,导致叶绿体结构的变化和对植物膜系统的损害,因而阐明光合速率下降的生理机制对于准确识别作物干旱程度和制定对策至关重要(Song et al.,2020). ...
作物水分利用效率研究方法及尺度传递研究进展
1
2019
... 水分利用效率(WUE)是植物基本的生理生态特征,高效的WUE有利于维持植物的碳水平衡和适应干旱环境(李民青等,2023).叶片尺度的WUE由光合作用与蒸腾作用共同决定,因而影响作物这两个生理过程的因素均会引起叶片WUE的变化(姜寒冰等,2019).本研究中,随着干旱胁迫的增加,玉米叶片的Pn和Tr均逐步降低,WUE最终表现为中旱与对照无显著差异,轻旱和重旱显著降低,10:00测量的叶水势,在轻旱与对照下无显著差异,中、重旱下显著降低.叶水势能够直接反映作物的水分亏缺情况,叶水势对WUE的影响是通过影响气孔行为实现的,当干旱胁迫刚开始时叶片可通过气孔的部分关闭来减少水分丧失,从而使叶水势在一定范围内保持恒定(张文丽等,2006),因而叶水势在轻旱处理下与对照处理无明显差异.但叶水势发生轻微下降都将提高植物激素脱落酸(Abscisic Acid,ABA)浓度,引起叶片气孔的轻微关闭以防止蒸腾性水分流失(Laxa et al.,2019),即气孔导度降低在叶片水势还没有明显变化之前就已经发生,这种生存策略会减少叶片CO2的吸收量,最终导致净光合速率下降,从而轻旱处理下WUE小于对照处理.当植物遭受较强的干旱胁迫后,会优先考虑生存而非生产力,而角质层损失的水量决定其存活能力(Mukarram et al.,2021),气孔的关闭不仅为了防止植物失水过度,同时可以使叶片水分利用效率最大化(战吉宬等,2005).叶水势在中旱下显著减小,气孔进一步关闭,以水分损失最小化为主要目的使蒸腾速率急剧降低(赵福年等,2019),此时蒸腾对干旱的响应比光合反应更强烈,因此蒸腾速率下降幅度要快于光合速率,也正是光合作用和蒸腾速率间的耦合关系促使中旱胁迫下水分利用效率最高,即气孔导度对叶片在碳同化和水分耗散的调节中符合最优控制的原则,这是植物抗旱策略的重要组成部分.重旱胁迫会导致叶绿素功能障碍并破坏光合器官(Cai et al.,2020),气孔对叶水势失去调节能力,因而重旱下叶水势降到极低,WUE显著减小,这与蔡福等(2017)的研究一致. ...
干旱-复水联动对东北春玉米光合生理与产量的影响
1
2023
... 干旱是农作物生长的主要制约因素之一(Lobell et al.,2014),玉米作为中国种植范围最广的谷类作物,在农业及国民经济中占据着重要地位(宋贺等,2019).全球气候变化背景下水热格局的改变,加剧了干旱等非生物胁迫因子对作物生长和产量的限制作用(李建查等,2018).近年来,光照、水分胁迫等环境因子对作物光合生理特征的影响得到了广泛关注(张仁和等,2011;Sharma and Zheng,2019;姜鹏等,2023),不同强度干旱过程对玉米生长发育、光合生理的影响也有较多研究,主要集中在拔节期和抽雄期(于文颖等,2015;宋贺等,2019;杨阳等,2022).萌发期和苗期(前期干旱)是春玉米干旱发生频率最高的阶段(任宗悦等,2020),特别是玉米营养生长阶段的七叶期(齐月等,2023),此时遭遇干旱会严重抑制春玉米的生长发育(卜令铎等,2010;Song et al.,2019).然而,目前针对干旱胁迫下春玉米七叶期的光合响应特征及其影响机制的研究尚显不足,因此本文通过研究春玉米七叶期光合参数变化特征及其对干旱的响应,试图明晰不同干旱程度下春玉米七叶期的叶片生理抗旱机制,以期为农业生产过程中春玉米生长前期的水分管理提供理论依据. ...
... 光合作用是地球生态系统一切生命活动的能量来源,也是作物产量形成的生理基础,其对环境变化十分敏感(卜令铎等,2010;李建查等,2018).作为植物最常见的生长逆境,干旱是抑制玉米生长和光合作用下降的重要原因(张仁和等,2011),限制着植物的光能利用率,甚至造成叶片结构的损伤(Mukarram et al.,2021),最终导致植物生物量及产量的下降(Jia et al.,2020).叶片光合作用对光的响应曲线是研究作物光合作用对环境变化响应的重要手段(李义博等,2017),可反映植物对不同光强的利用规律,并指示植物的生长情况及其抗逆性.通常以气孔导度、净光合速率、蒸腾速率及水分利用效率等指标来表征作物的光合特性(于文颖等,2015).气孔不仅是水分和光合原料进入细胞的关键通道,在干旱感知和信号传导过程中也发挥着不可或缺的作用(Laxa et al.,2019).Hetherington和Woodward(2003)研究指出,气孔面积仅占叶片面积的5%,但经由气孔蒸发的水分却高达70%.由此可见,气孔调节作为植物抵御胁迫环境的核心机制,对于维持植物体内水分平衡、保障光合作用正常进行意义重大.气孔导度作为衡量气孔变化特征的重要指标,其大小直接影响叶片中CO2的交换速率和水分的蒸腾速率(龚容和高琼,2015).适度水分亏缺下叶片可通过改变气孔的形态和气孔导度的大小来控制叶片内部CO2浓度和蒸腾速率,协调碳同化和水分消耗之间的关系(Ennahli and Earl,2005),同时影响叶片水分利用效率(Water Use Efficiency,WUE),最终形成高效的抗旱策略以适应干旱环境(李民青等,2023),随着干旱条件的进一步发展,作物生产可能出现环境约束向生理生态约束的转变,植物气孔可能关闭,导致叶绿体结构的变化和对植物膜系统的损害,因而阐明光合速率下降的生理机制对于准确识别作物干旱程度和制定对策至关重要(Song et al.,2020). ...
... 干旱是农作物生长的主要制约因素之一(Lobell et al.,2014),玉米作为中国种植范围最广的谷类作物,在农业及国民经济中占据着重要地位(宋贺等,2019).全球气候变化背景下水热格局的改变,加剧了干旱等非生物胁迫因子对作物生长和产量的限制作用(李建查等,2018).近年来,光照、水分胁迫等环境因子对作物光合生理特征的影响得到了广泛关注(张仁和等,2011;Sharma and Zheng,2019;姜鹏等,2023),不同强度干旱过程对玉米生长发育、光合生理的影响也有较多研究,主要集中在拔节期和抽雄期(于文颖等,2015;宋贺等,2019;杨阳等,2022).萌发期和苗期(前期干旱)是春玉米干旱发生频率最高的阶段(任宗悦等,2020),特别是玉米营养生长阶段的七叶期(齐月等,2023),此时遭遇干旱会严重抑制春玉米的生长发育(卜令铎等,2010;Song et al.,2019).然而,目前针对干旱胁迫下春玉米七叶期的光合响应特征及其影响机制的研究尚显不足,因此本文通过研究春玉米七叶期光合参数变化特征及其对干旱的响应,试图明晰不同干旱程度下春玉米七叶期的叶片生理抗旱机制,以期为农业生产过程中春玉米生长前期的水分管理提供理论依据. ...
7种荒漠木本植物枝干与叶片光合特征及其影响因素
2
2023
... 光合作用是地球生态系统一切生命活动的能量来源,也是作物产量形成的生理基础,其对环境变化十分敏感(卜令铎等,2010;李建查等,2018).作为植物最常见的生长逆境,干旱是抑制玉米生长和光合作用下降的重要原因(张仁和等,2011),限制着植物的光能利用率,甚至造成叶片结构的损伤(Mukarram et al.,2021),最终导致植物生物量及产量的下降(Jia et al.,2020).叶片光合作用对光的响应曲线是研究作物光合作用对环境变化响应的重要手段(李义博等,2017),可反映植物对不同光强的利用规律,并指示植物的生长情况及其抗逆性.通常以气孔导度、净光合速率、蒸腾速率及水分利用效率等指标来表征作物的光合特性(于文颖等,2015).气孔不仅是水分和光合原料进入细胞的关键通道,在干旱感知和信号传导过程中也发挥着不可或缺的作用(Laxa et al.,2019).Hetherington和Woodward(2003)研究指出,气孔面积仅占叶片面积的5%,但经由气孔蒸发的水分却高达70%.由此可见,气孔调节作为植物抵御胁迫环境的核心机制,对于维持植物体内水分平衡、保障光合作用正常进行意义重大.气孔导度作为衡量气孔变化特征的重要指标,其大小直接影响叶片中CO2的交换速率和水分的蒸腾速率(龚容和高琼,2015).适度水分亏缺下叶片可通过改变气孔的形态和气孔导度的大小来控制叶片内部CO2浓度和蒸腾速率,协调碳同化和水分消耗之间的关系(Ennahli and Earl,2005),同时影响叶片水分利用效率(Water Use Efficiency,WUE),最终形成高效的抗旱策略以适应干旱环境(李民青等,2023),随着干旱条件的进一步发展,作物生产可能出现环境约束向生理生态约束的转变,植物气孔可能关闭,导致叶绿体结构的变化和对植物膜系统的损害,因而阐明光合速率下降的生理机制对于准确识别作物干旱程度和制定对策至关重要(Song et al.,2020). ...
... 水分利用效率(WUE)是植物基本的生理生态特征,高效的WUE有利于维持植物的碳水平衡和适应干旱环境(李民青等,2023).叶片尺度的WUE由光合作用与蒸腾作用共同决定,因而影响作物这两个生理过程的因素均会引起叶片WUE的变化(姜寒冰等,2019).本研究中,随着干旱胁迫的增加,玉米叶片的Pn和Tr均逐步降低,WUE最终表现为中旱与对照无显著差异,轻旱和重旱显著降低,10:00测量的叶水势,在轻旱与对照下无显著差异,中、重旱下显著降低.叶水势能够直接反映作物的水分亏缺情况,叶水势对WUE的影响是通过影响气孔行为实现的,当干旱胁迫刚开始时叶片可通过气孔的部分关闭来减少水分丧失,从而使叶水势在一定范围内保持恒定(张文丽等,2006),因而叶水势在轻旱处理下与对照处理无明显差异.但叶水势发生轻微下降都将提高植物激素脱落酸(Abscisic Acid,ABA)浓度,引起叶片气孔的轻微关闭以防止蒸腾性水分流失(Laxa et al.,2019),即气孔导度降低在叶片水势还没有明显变化之前就已经发生,这种生存策略会减少叶片CO2的吸收量,最终导致净光合速率下降,从而轻旱处理下WUE小于对照处理.当植物遭受较强的干旱胁迫后,会优先考虑生存而非生产力,而角质层损失的水量决定其存活能力(Mukarram et al.,2021),气孔的关闭不仅为了防止植物失水过度,同时可以使叶片水分利用效率最大化(战吉宬等,2005).叶水势在中旱下显著减小,气孔进一步关闭,以水分损失最小化为主要目的使蒸腾速率急剧降低(赵福年等,2019),此时蒸腾对干旱的响应比光合反应更强烈,因此蒸腾速率下降幅度要快于光合速率,也正是光合作用和蒸腾速率间的耦合关系促使中旱胁迫下水分利用效率最高,即气孔导度对叶片在碳同化和水分耗散的调节中符合最优控制的原则,这是植物抗旱策略的重要组成部分.重旱胁迫会导致叶绿素功能障碍并破坏光合器官(Cai et al.,2020),气孔对叶水势失去调节能力,因而重旱下叶水势降到极低,WUE显著减小,这与蔡福等(2017)的研究一致. ...
C4植物玉米的光合-光响应曲线模拟研究
4
2017
... 光合作用是地球生态系统一切生命活动的能量来源,也是作物产量形成的生理基础,其对环境变化十分敏感(卜令铎等,2010;李建查等,2018).作为植物最常见的生长逆境,干旱是抑制玉米生长和光合作用下降的重要原因(张仁和等,2011),限制着植物的光能利用率,甚至造成叶片结构的损伤(Mukarram et al.,2021),最终导致植物生物量及产量的下降(Jia et al.,2020).叶片光合作用对光的响应曲线是研究作物光合作用对环境变化响应的重要手段(李义博等,2017),可反映植物对不同光强的利用规律,并指示植物的生长情况及其抗逆性.通常以气孔导度、净光合速率、蒸腾速率及水分利用效率等指标来表征作物的光合特性(于文颖等,2015).气孔不仅是水分和光合原料进入细胞的关键通道,在干旱感知和信号传导过程中也发挥着不可或缺的作用(Laxa et al.,2019).Hetherington和Woodward(2003)研究指出,气孔面积仅占叶片面积的5%,但经由气孔蒸发的水分却高达70%.由此可见,气孔调节作为植物抵御胁迫环境的核心机制,对于维持植物体内水分平衡、保障光合作用正常进行意义重大.气孔导度作为衡量气孔变化特征的重要指标,其大小直接影响叶片中CO2的交换速率和水分的蒸腾速率(龚容和高琼,2015).适度水分亏缺下叶片可通过改变气孔的形态和气孔导度的大小来控制叶片内部CO2浓度和蒸腾速率,协调碳同化和水分消耗之间的关系(Ennahli and Earl,2005),同时影响叶片水分利用效率(Water Use Efficiency,WUE),最终形成高效的抗旱策略以适应干旱环境(李民青等,2023),随着干旱条件的进一步发展,作物生产可能出现环境约束向生理生态约束的转变,植物气孔可能关闭,导致叶绿体结构的变化和对植物膜系统的损害,因而阐明光合速率下降的生理机制对于准确识别作物干旱程度和制定对策至关重要(Song et al.,2020). ...
... 植物光合作用主要受气孔和非气孔因素影响(Jia et al.,2020),前者指干旱胁迫导致的气孔导度降低,CO2进入叶片的通道受阻而使光合速率下降(Song et al.,2020),后者主要是叶肉细胞电子传递和光合磷酸化受阻,暗反应中相关酶活性降低,导致光合速率下降(Farquhar and Sharkey,1982).宋丰萍和蒙祖庆(2018)研究表明,随着干旱胁迫的逐步加剧、持续时间不断延长,气孔和非气孔因素往往会同时对植物光合作用产生影响,在这一过程中,非气孔因素最终会成为主导限制因子.基于此,麻雪艳和周广胜(2018)提出,气孔限制向非气孔限制的转折点,可作为判定玉米是否受到严重干旱胁迫的重要依据. ...
干旱胁迫下春玉米叶片光合参数对叶温的响应
2
2023
... 干旱是农作物生长的主要制约因素之一(Lobell et al.,2014),玉米作为中国种植范围最广的谷类作物,在农业及国民经济中占据着重要地位(宋贺等,2019).全球气候变化背景下水热格局的改变,加剧了干旱等非生物胁迫因子对作物生长和产量的限制作用(李建查等,2018).近年来,光照、水分胁迫等环境因子对作物光合生理特征的影响得到了广泛关注(张仁和等,2011;Sharma and Zheng,2019;姜鹏等,2023),不同强度干旱过程对玉米生长发育、光合生理的影响也有较多研究,主要集中在拔节期和抽雄期(于文颖等,2015;宋贺等,2019;杨阳等,2022).萌发期和苗期(前期干旱)是春玉米干旱发生频率最高的阶段(任宗悦等,2020),特别是玉米营养生长阶段的七叶期(齐月等,2023),此时遭遇干旱会严重抑制春玉米的生长发育(卜令铎等,2010;Song et al.,2019).然而,目前针对干旱胁迫下春玉米七叶期的光合响应特征及其影响机制的研究尚显不足,因此本文通过研究春玉米七叶期光合参数变化特征及其对干旱的响应,试图明晰不同干旱程度下春玉米七叶期的叶片生理抗旱机制,以期为农业生产过程中春玉米生长前期的水分管理提供理论依据. ...
... 干旱是农作物生长的主要制约因素之一(Lobell et al.,2014),玉米作为中国种植范围最广的谷类作物,在农业及国民经济中占据着重要地位(宋贺等,2019).全球气候变化背景下水热格局的改变,加剧了干旱等非生物胁迫因子对作物生长和产量的限制作用(李建查等,2018).近年来,光照、水分胁迫等环境因子对作物光合生理特征的影响得到了广泛关注(张仁和等,2011;Sharma and Zheng,2019;姜鹏等,2023),不同强度干旱过程对玉米生长发育、光合生理的影响也有较多研究,主要集中在拔节期和抽雄期(于文颖等,2015;宋贺等,2019;杨阳等,2022).萌发期和苗期(前期干旱)是春玉米干旱发生频率最高的阶段(任宗悦等,2020),特别是玉米营养生长阶段的七叶期(齐月等,2023),此时遭遇干旱会严重抑制春玉米的生长发育(卜令铎等,2010;Song et al.,2019).然而,目前针对干旱胁迫下春玉米七叶期的光合响应特征及其影响机制的研究尚显不足,因此本文通过研究春玉米七叶期光合参数变化特征及其对干旱的响应,试图明晰不同干旱程度下春玉米七叶期的叶片生理抗旱机制,以期为农业生产过程中春玉米生长前期的水分管理提供理论依据. ...
干旱胁迫下作物光合参数研究进展
1
2018
... 植物光合作用主要受气孔和非气孔因素影响(Jia et al.,2020),前者指干旱胁迫导致的气孔导度降低,CO2进入叶片的通道受阻而使光合速率下降(Song et al.,2020),后者主要是叶肉细胞电子传递和光合磷酸化受阻,暗反应中相关酶活性降低,导致光合速率下降(Farquhar and Sharkey,1982).宋丰萍和蒙祖庆(2018)研究表明,随着干旱胁迫的逐步加剧、持续时间不断延长,气孔和非气孔因素往往会同时对植物光合作用产生影响,在这一过程中,非气孔因素最终会成为主导限制因子.基于此,麻雪艳和周广胜(2018)提出,气孔限制向非气孔限制的转折点,可作为判定玉米是否受到严重干旱胁迫的重要依据. ...
玉米光合生理参数对全生育期干旱与拔节后干旱过程的响应
3
2019
... 干旱是农作物生长的主要制约因素之一(Lobell et al.,2014),玉米作为中国种植范围最广的谷类作物,在农业及国民经济中占据着重要地位(宋贺等,2019).全球气候变化背景下水热格局的改变,加剧了干旱等非生物胁迫因子对作物生长和产量的限制作用(李建查等,2018).近年来,光照、水分胁迫等环境因子对作物光合生理特征的影响得到了广泛关注(张仁和等,2011;Sharma and Zheng,2019;姜鹏等,2023),不同强度干旱过程对玉米生长发育、光合生理的影响也有较多研究,主要集中在拔节期和抽雄期(于文颖等,2015;宋贺等,2019;杨阳等,2022).萌发期和苗期(前期干旱)是春玉米干旱发生频率最高的阶段(任宗悦等,2020),特别是玉米营养生长阶段的七叶期(齐月等,2023),此时遭遇干旱会严重抑制春玉米的生长发育(卜令铎等,2010;Song et al.,2019).然而,目前针对干旱胁迫下春玉米七叶期的光合响应特征及其影响机制的研究尚显不足,因此本文通过研究春玉米七叶期光合参数变化特征及其对干旱的响应,试图明晰不同干旱程度下春玉米七叶期的叶片生理抗旱机制,以期为农业生产过程中春玉米生长前期的水分管理提供理论依据. ...
... ;宋贺等,2019;杨阳等,2022).萌发期和苗期(前期干旱)是春玉米干旱发生频率最高的阶段(任宗悦等,2020),特别是玉米营养生长阶段的七叶期(齐月等,2023),此时遭遇干旱会严重抑制春玉米的生长发育(卜令铎等,2010;Song et al.,2019).然而,目前针对干旱胁迫下春玉米七叶期的光合响应特征及其影响机制的研究尚显不足,因此本文通过研究春玉米七叶期光合参数变化特征及其对干旱的响应,试图明晰不同干旱程度下春玉米七叶期的叶片生理抗旱机制,以期为农业生产过程中春玉米生长前期的水分管理提供理论依据. ...
... 植物受到干旱胁迫时光合作用被限制,碳同化能力下降(Neto et al.,2017),光合速率的下降由气孔和非气孔因素共同决定(Jia et al.,2020).叶子飘等(2019)研究表明,当所有气孔开闭行为保持一致时,判断叶片光合速率下降是否由气孔因素主导,需依据胞间CO2浓度(Ci)和气孔限制值的变化趋势来判定,而非单纯关注Ci的下降幅度.根据Farquhar和Sharkey(1982)的观点,在Pn减小的情况下,Ci下降且Ls升高时,气孔限制占主导作用,当Ci升高且Ls下降时,非气孔因素占主导作用.本研究中,随着光照增强,轻旱胁迫下玉米Gs相较于对照处理显著下降,叶片Ci增加,Ls减小,根据Ci的变化方向判断此时以非气孔限制为主导因素使净光合速率降低.但许大全(2013)指出叶片气孔的不均关闭现象会使Ci恒定不变,水分胁迫条件下容易造成非气孔限制的假象,当作物突然遭受水分胁迫,植物为维持正常的叶水势,通过关闭部分气孔的方式减少水分蒸腾(Sharma and Zheng,2019),计算的Ci会高于或者等于实际值,进而造成非气孔限制的假象.Medrano等(2002)提出,可由光饱和的气孔导度作为干旱条件下光合作用的气孔和非气孔限制之间的拐点指标,最大气孔导度值在0.05 mol·m-2·s-1左右时即为拐点值,因而轻旱下主要是气孔限制影响了叶片碳同化能力,导致光合速率下降.中旱处理下玉米叶片Ci减小,Ls增加,Gs和Pn均显著下降,结合最大气孔导度拐点值,此时叶片净光合速率下降的因素以气孔限制为主导,非气孔限制逐渐增强.中旱处理下植物气孔的开放程度进一步降低,气孔阻力增加,限制了光合作用原料的供应,导致Ci降低(Song et al.,2020),最终使作物光合作用速率降低,叶片通过气孔调节的方式降低水分蒸腾,避免干旱对叶片结构的损伤,这可能是本文中旱处理下气孔限制始终占主导因素的原因,体现了植物适应胁迫环境的气孔调节机制,表明玉米属较耐旱的作物,在受到干旱胁迫后作物光合系统的自身调节能力较强,可忍耐一定程度的水分亏缺(陈图强等,2023).发生轻微干旱时,植物的抗氧化防御机制一般可以及时清除活性氧自由基以避免膜损伤(Jia et al.,2020),但重旱下土壤水分胁迫超出了植物可应对的阈值,叶片不能得到生存所需的最低水分,植物收支平衡被打破,叶肉细胞光合活性降低,抗氧化防御机理极其脆弱,活性氧的积累会破坏光系统(Sharma and Zheng,2019),植物无法维持正常生理活动.重旱下玉米叶片Ci显著增加,Ls明显减小,非气孔因素限制作物的光合作用,因而净光合速率等光合参数趋近于零. ...
... 干旱是农作物生长的主要制约因素之一(Lobell et al.,2014),玉米作为中国种植范围最广的谷类作物,在农业及国民经济中占据着重要地位(宋贺等,2019).全球气候变化背景下水热格局的改变,加剧了干旱等非生物胁迫因子对作物生长和产量的限制作用(李建查等,2018).近年来,光照、水分胁迫等环境因子对作物光合生理特征的影响得到了广泛关注(张仁和等,2011;Sharma and Zheng,2019;姜鹏等,2023),不同强度干旱过程对玉米生长发育、光合生理的影响也有较多研究,主要集中在拔节期和抽雄期(于文颖等,2015;宋贺等,2019;杨阳等,2022).萌发期和苗期(前期干旱)是春玉米干旱发生频率最高的阶段(任宗悦等,2020),特别是玉米营养生长阶段的七叶期(齐月等,2023),此时遭遇干旱会严重抑制春玉米的生长发育(卜令铎等,2010;Song et al.,2019).然而,目前针对干旱胁迫下春玉米七叶期的光合响应特征及其影响机制的研究尚显不足,因此本文通过研究春玉米七叶期光合参数变化特征及其对干旱的响应,试图明晰不同干旱程度下春玉米七叶期的叶片生理抗旱机制,以期为农业生产过程中春玉米生长前期的水分管理提供理论依据. ...
... 植物受到干旱胁迫时光合作用被限制,碳同化能力下降(Neto et al.,2017),光合速率的下降由气孔和非气孔因素共同决定(Jia et al.,2020).叶子飘等(2019)研究表明,当所有气孔开闭行为保持一致时,判断叶片光合速率下降是否由气孔因素主导,需依据胞间CO2浓度(Ci)和气孔限制值的变化趋势来判定,而非单纯关注Ci的下降幅度.根据Farquhar和Sharkey(1982)的观点,在Pn减小的情况下,Ci下降且Ls升高时,气孔限制占主导作用,当Ci升高且Ls下降时,非气孔因素占主导作用.本研究中,随着光照增强,轻旱胁迫下玉米Gs相较于对照处理显著下降,叶片Ci增加,Ls减小,根据Ci的变化方向判断此时以非气孔限制为主导因素使净光合速率降低.但许大全(2013)指出叶片气孔的不均关闭现象会使Ci恒定不变,水分胁迫条件下容易造成非气孔限制的假象,当作物突然遭受水分胁迫,植物为维持正常的叶水势,通过关闭部分气孔的方式减少水分蒸腾(Sharma and Zheng,2019),计算的Ci会高于或者等于实际值,进而造成非气孔限制的假象.Medrano等(2002)提出,可由光饱和的气孔导度作为干旱条件下光合作用的气孔和非气孔限制之间的拐点指标,最大气孔导度值在0.05 mol·m-2·s-1左右时即为拐点值,因而轻旱下主要是气孔限制影响了叶片碳同化能力,导致光合速率下降.中旱处理下玉米叶片Ci减小,Ls增加,Gs和Pn均显著下降,结合最大气孔导度拐点值,此时叶片净光合速率下降的因素以气孔限制为主导,非气孔限制逐渐增强.中旱处理下植物气孔的开放程度进一步降低,气孔阻力增加,限制了光合作用原料的供应,导致Ci降低(Song et al.,2020),最终使作物光合作用速率降低,叶片通过气孔调节的方式降低水分蒸腾,避免干旱对叶片结构的损伤,这可能是本文中旱处理下气孔限制始终占主导因素的原因,体现了植物适应胁迫环境的气孔调节机制,表明玉米属较耐旱的作物,在受到干旱胁迫后作物光合系统的自身调节能力较强,可忍耐一定程度的水分亏缺(陈图强等,2023).发生轻微干旱时,植物的抗氧化防御机制一般可以及时清除活性氧自由基以避免膜损伤(Jia et al.,2020),但重旱下土壤水分胁迫超出了植物可应对的阈值,叶片不能得到生存所需的最低水分,植物收支平衡被打破,叶肉细胞光合活性降低,抗氧化防御机理极其脆弱,活性氧的积累会破坏光系统(Sharma and Zheng,2019),植物无法维持正常生理活动.重旱下玉米叶片Ci显著增加,Ls明显减小,非气孔因素限制作物的光合作用,因而净光合速率等光合参数趋近于零. ...
不同生育期玉米叶片光合特性及水分利用效率对水分胁迫的响应
2
2015
... 光合作用是地球生态系统一切生命活动的能量来源,也是作物产量形成的生理基础,其对环境变化十分敏感(卜令铎等,2010;李建查等,2018).作为植物最常见的生长逆境,干旱是抑制玉米生长和光合作用下降的重要原因(张仁和等,2011),限制着植物的光能利用率,甚至造成叶片结构的损伤(Mukarram et al.,2021),最终导致植物生物量及产量的下降(Jia et al.,2020).叶片光合作用对光的响应曲线是研究作物光合作用对环境变化响应的重要手段(李义博等,2017),可反映植物对不同光强的利用规律,并指示植物的生长情况及其抗逆性.通常以气孔导度、净光合速率、蒸腾速率及水分利用效率等指标来表征作物的光合特性(于文颖等,2015).气孔不仅是水分和光合原料进入细胞的关键通道,在干旱感知和信号传导过程中也发挥着不可或缺的作用(Laxa et al.,2019).Hetherington和Woodward(2003)研究指出,气孔面积仅占叶片面积的5%,但经由气孔蒸发的水分却高达70%.由此可见,气孔调节作为植物抵御胁迫环境的核心机制,对于维持植物体内水分平衡、保障光合作用正常进行意义重大.气孔导度作为衡量气孔变化特征的重要指标,其大小直接影响叶片中CO2的交换速率和水分的蒸腾速率(龚容和高琼,2015).适度水分亏缺下叶片可通过改变气孔的形态和气孔导度的大小来控制叶片内部CO2浓度和蒸腾速率,协调碳同化和水分消耗之间的关系(Ennahli and Earl,2005),同时影响叶片水分利用效率(Water Use Efficiency,WUE),最终形成高效的抗旱策略以适应干旱环境(李民青等,2023),随着干旱条件的进一步发展,作物生产可能出现环境约束向生理生态约束的转变,植物气孔可能关闭,导致叶绿体结构的变化和对植物膜系统的损害,因而阐明光合速率下降的生理机制对于准确识别作物干旱程度和制定对策至关重要(Song et al.,2020). ...
... 干旱是农作物生长的主要制约因素之一(Lobell et al.,2014),玉米作为中国种植范围最广的谷类作物,在农业及国民经济中占据着重要地位(宋贺等,2019).全球气候变化背景下水热格局的改变,加剧了干旱等非生物胁迫因子对作物生长和产量的限制作用(李建查等,2018).近年来,光照、水分胁迫等环境因子对作物光合生理特征的影响得到了广泛关注(张仁和等,2011;Sharma and Zheng,2019;姜鹏等,2023),不同强度干旱过程对玉米生长发育、光合生理的影响也有较多研究,主要集中在拔节期和抽雄期(于文颖等,2015;宋贺等,2019;杨阳等,2022).萌发期和苗期(前期干旱)是春玉米干旱发生频率最高的阶段(任宗悦等,2020),特别是玉米营养生长阶段的七叶期(齐月等,2023),此时遭遇干旱会严重抑制春玉米的生长发育(卜令铎等,2010;Song et al.,2019).然而,目前针对干旱胁迫下春玉米七叶期的光合响应特征及其影响机制的研究尚显不足,因此本文通过研究春玉米七叶期光合参数变化特征及其对干旱的响应,试图明晰不同干旱程度下春玉米七叶期的叶片生理抗旱机制,以期为农业生产过程中春玉米生长前期的水分管理提供理论依据. ...
弱光下生长的葡萄叶片蒸腾速率和气孔结构的变化
1
2005
... 水分利用效率(WUE)是植物基本的生理生态特征,高效的WUE有利于维持植物的碳水平衡和适应干旱环境(李民青等,2023).叶片尺度的WUE由光合作用与蒸腾作用共同决定,因而影响作物这两个生理过程的因素均会引起叶片WUE的变化(姜寒冰等,2019).本研究中,随着干旱胁迫的增加,玉米叶片的Pn和Tr均逐步降低,WUE最终表现为中旱与对照无显著差异,轻旱和重旱显著降低,10:00测量的叶水势,在轻旱与对照下无显著差异,中、重旱下显著降低.叶水势能够直接反映作物的水分亏缺情况,叶水势对WUE的影响是通过影响气孔行为实现的,当干旱胁迫刚开始时叶片可通过气孔的部分关闭来减少水分丧失,从而使叶水势在一定范围内保持恒定(张文丽等,2006),因而叶水势在轻旱处理下与对照处理无明显差异.但叶水势发生轻微下降都将提高植物激素脱落酸(Abscisic Acid,ABA)浓度,引起叶片气孔的轻微关闭以防止蒸腾性水分流失(Laxa et al.,2019),即气孔导度降低在叶片水势还没有明显变化之前就已经发生,这种生存策略会减少叶片CO2的吸收量,最终导致净光合速率下降,从而轻旱处理下WUE小于对照处理.当植物遭受较强的干旱胁迫后,会优先考虑生存而非生产力,而角质层损失的水量决定其存活能力(Mukarram et al.,2021),气孔的关闭不仅为了防止植物失水过度,同时可以使叶片水分利用效率最大化(战吉宬等,2005).叶水势在中旱下显著减小,气孔进一步关闭,以水分损失最小化为主要目的使蒸腾速率急剧降低(赵福年等,2019),此时蒸腾对干旱的响应比光合反应更强烈,因此蒸腾速率下降幅度要快于光合速率,也正是光合作用和蒸腾速率间的耦合关系促使中旱胁迫下水分利用效率最高,即气孔导度对叶片在碳同化和水分耗散的调节中符合最优控制的原则,这是植物抗旱策略的重要组成部分.重旱胁迫会导致叶绿素功能障碍并破坏光合器官(Cai et al.,2020),气孔对叶水势失去调节能力,因而重旱下叶水势降到极低,WUE显著减小,这与蔡福等(2017)的研究一致. ...
干旱胁迫对甘肃中部春小麦生理性状及灌水利用效率的影响
1
2019
... 于5月30日春玉米七叶期开始时进行控水(即播种后第30天),随后开展各项观测工作.每天定时观测土壤水分、光合生理参数及叶水势等指标,直至植株萎蔫死亡,试验结束.在整个试验时段内,每日06:00和10:00采集第一片完全展开叶,使用WP4露点水势仪测定叶水势,每个处理设置3次重复(张健等,2019).在试验时段内,为确保测量数据的准确性与稳定性,选定每天上午09:00—11:00,利用Li-6400XT光合测量系统(LI-COR,Lincoln,USA)测定春玉米叶片的光合生理指标.选取玉米顶部第一片展开叶为测量对象,测量时指定使用人工红蓝光源,CO2浓度采用小钢瓶控制,样品室浓度设置为400 μmol·mol-1,叶室温度设置为25 ℃,在1 800 μmol·m-2·s-1的光强下充分诱导,待光合参数稳定后,利用自动测量程序进行测量,光合有效辐射(Photosynthetically Active Radiation,PAR)梯度设定为2 400、2 100、1 800、1 500、1 200、1 000、800、600、400、200、150、100、75、50、25、0 μmol·m-2·s-1,每个处理3次重复.测定内容包括:净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs),并计算气孔限制值(Ls)(Farquhar and Sharkey,1982)和瞬时水分利用效率(WUE),具体公式如下: ...
干旱胁迫对玉米苗期叶片光合作用和保护酶的影响
2
2011
... 光合作用是地球生态系统一切生命活动的能量来源,也是作物产量形成的生理基础,其对环境变化十分敏感(卜令铎等,2010;李建查等,2018).作为植物最常见的生长逆境,干旱是抑制玉米生长和光合作用下降的重要原因(张仁和等,2011),限制着植物的光能利用率,甚至造成叶片结构的损伤(Mukarram et al.,2021),最终导致植物生物量及产量的下降(Jia et al.,2020).叶片光合作用对光的响应曲线是研究作物光合作用对环境变化响应的重要手段(李义博等,2017),可反映植物对不同光强的利用规律,并指示植物的生长情况及其抗逆性.通常以气孔导度、净光合速率、蒸腾速率及水分利用效率等指标来表征作物的光合特性(于文颖等,2015).气孔不仅是水分和光合原料进入细胞的关键通道,在干旱感知和信号传导过程中也发挥着不可或缺的作用(Laxa et al.,2019).Hetherington和Woodward(2003)研究指出,气孔面积仅占叶片面积的5%,但经由气孔蒸发的水分却高达70%.由此可见,气孔调节作为植物抵御胁迫环境的核心机制,对于维持植物体内水分平衡、保障光合作用正常进行意义重大.气孔导度作为衡量气孔变化特征的重要指标,其大小直接影响叶片中CO2的交换速率和水分的蒸腾速率(龚容和高琼,2015).适度水分亏缺下叶片可通过改变气孔的形态和气孔导度的大小来控制叶片内部CO2浓度和蒸腾速率,协调碳同化和水分消耗之间的关系(Ennahli and Earl,2005),同时影响叶片水分利用效率(Water Use Efficiency,WUE),最终形成高效的抗旱策略以适应干旱环境(李民青等,2023),随着干旱条件的进一步发展,作物生产可能出现环境约束向生理生态约束的转变,植物气孔可能关闭,导致叶绿体结构的变化和对植物膜系统的损害,因而阐明光合速率下降的生理机制对于准确识别作物干旱程度和制定对策至关重要(Song et al.,2020). ...
... 干旱是农作物生长的主要制约因素之一(Lobell et al.,2014),玉米作为中国种植范围最广的谷类作物,在农业及国民经济中占据着重要地位(宋贺等,2019).全球气候变化背景下水热格局的改变,加剧了干旱等非生物胁迫因子对作物生长和产量的限制作用(李建查等,2018).近年来,光照、水分胁迫等环境因子对作物光合生理特征的影响得到了广泛关注(张仁和等,2011;Sharma and Zheng,2019;姜鹏等,2023),不同强度干旱过程对玉米生长发育、光合生理的影响也有较多研究,主要集中在拔节期和抽雄期(于文颖等,2015;宋贺等,2019;杨阳等,2022).萌发期和苗期(前期干旱)是春玉米干旱发生频率最高的阶段(任宗悦等,2020),特别是玉米营养生长阶段的七叶期(齐月等,2023),此时遭遇干旱会严重抑制春玉米的生长发育(卜令铎等,2010;Song et al.,2019).然而,目前针对干旱胁迫下春玉米七叶期的光合响应特征及其影响机制的研究尚显不足,因此本文通过研究春玉米七叶期光合参数变化特征及其对干旱的响应,试图明晰不同干旱程度下春玉米七叶期的叶片生理抗旱机制,以期为农业生产过程中春玉米生长前期的水分管理提供理论依据. ...
土壤逐渐干旱下玉米幼苗光合速率与蒸腾速率变化的研究
1
2006
... 水分利用效率(WUE)是植物基本的生理生态特征,高效的WUE有利于维持植物的碳水平衡和适应干旱环境(李民青等,2023).叶片尺度的WUE由光合作用与蒸腾作用共同决定,因而影响作物这两个生理过程的因素均会引起叶片WUE的变化(姜寒冰等,2019).本研究中,随着干旱胁迫的增加,玉米叶片的Pn和Tr均逐步降低,WUE最终表现为中旱与对照无显著差异,轻旱和重旱显著降低,10:00测量的叶水势,在轻旱与对照下无显著差异,中、重旱下显著降低.叶水势能够直接反映作物的水分亏缺情况,叶水势对WUE的影响是通过影响气孔行为实现的,当干旱胁迫刚开始时叶片可通过气孔的部分关闭来减少水分丧失,从而使叶水势在一定范围内保持恒定(张文丽等,2006),因而叶水势在轻旱处理下与对照处理无明显差异.但叶水势发生轻微下降都将提高植物激素脱落酸(Abscisic Acid,ABA)浓度,引起叶片气孔的轻微关闭以防止蒸腾性水分流失(Laxa et al.,2019),即气孔导度降低在叶片水势还没有明显变化之前就已经发生,这种生存策略会减少叶片CO2的吸收量,最终导致净光合速率下降,从而轻旱处理下WUE小于对照处理.当植物遭受较强的干旱胁迫后,会优先考虑生存而非生产力,而角质层损失的水量决定其存活能力(Mukarram et al.,2021),气孔的关闭不仅为了防止植物失水过度,同时可以使叶片水分利用效率最大化(战吉宬等,2005).叶水势在中旱下显著减小,气孔进一步关闭,以水分损失最小化为主要目的使蒸腾速率急剧降低(赵福年等,2019),此时蒸腾对干旱的响应比光合反应更强烈,因此蒸腾速率下降幅度要快于光合速率,也正是光合作用和蒸腾速率间的耦合关系促使中旱胁迫下水分利用效率最高,即气孔导度对叶片在碳同化和水分耗散的调节中符合最优控制的原则,这是植物抗旱策略的重要组成部分.重旱胁迫会导致叶绿素功能障碍并破坏光合器官(Cai et al.,2020),气孔对叶水势失去调节能力,因而重旱下叶水势降到极低,WUE显著减小,这与蔡福等(2017)的研究一致. ...
作物内禀水分利用效率变化
1
2019
... 水分利用效率(WUE)是植物基本的生理生态特征,高效的WUE有利于维持植物的碳水平衡和适应干旱环境(李民青等,2023).叶片尺度的WUE由光合作用与蒸腾作用共同决定,因而影响作物这两个生理过程的因素均会引起叶片WUE的变化(姜寒冰等,2019).本研究中,随着干旱胁迫的增加,玉米叶片的Pn和Tr均逐步降低,WUE最终表现为中旱与对照无显著差异,轻旱和重旱显著降低,10:00测量的叶水势,在轻旱与对照下无显著差异,中、重旱下显著降低.叶水势能够直接反映作物的水分亏缺情况,叶水势对WUE的影响是通过影响气孔行为实现的,当干旱胁迫刚开始时叶片可通过气孔的部分关闭来减少水分丧失,从而使叶水势在一定范围内保持恒定(张文丽等,2006),因而叶水势在轻旱处理下与对照处理无明显差异.但叶水势发生轻微下降都将提高植物激素脱落酸(Abscisic Acid,ABA)浓度,引起叶片气孔的轻微关闭以防止蒸腾性水分流失(Laxa et al.,2019),即气孔导度降低在叶片水势还没有明显变化之前就已经发生,这种生存策略会减少叶片CO2的吸收量,最终导致净光合速率下降,从而轻旱处理下WUE小于对照处理.当植物遭受较强的干旱胁迫后,会优先考虑生存而非生产力,而角质层损失的水量决定其存活能力(Mukarram et al.,2021),气孔的关闭不仅为了防止植物失水过度,同时可以使叶片水分利用效率最大化(战吉宬等,2005).叶水势在中旱下显著减小,气孔进一步关闭,以水分损失最小化为主要目的使蒸腾速率急剧降低(赵福年等,2019),此时蒸腾对干旱的响应比光合反应更强烈,因此蒸腾速率下降幅度要快于光合速率,也正是光合作用和蒸腾速率间的耦合关系促使中旱胁迫下水分利用效率最高,即气孔导度对叶片在碳同化和水分耗散的调节中符合最优控制的原则,这是植物抗旱策略的重要组成部分.重旱胁迫会导致叶绿素功能障碍并破坏光合器官(Cai et al.,2020),气孔对叶水势失去调节能力,因而重旱下叶水势降到极低,WUE显著减小,这与蔡福等(2017)的研究一致. ...
Maize (Zea mays L.) physiological responses to drought and rewatering, and the associations with water stress degree
1
2020
... 水分利用效率(WUE)是植物基本的生理生态特征,高效的WUE有利于维持植物的碳水平衡和适应干旱环境(李民青等,2023).叶片尺度的WUE由光合作用与蒸腾作用共同决定,因而影响作物这两个生理过程的因素均会引起叶片WUE的变化(姜寒冰等,2019).本研究中,随着干旱胁迫的增加,玉米叶片的Pn和Tr均逐步降低,WUE最终表现为中旱与对照无显著差异,轻旱和重旱显著降低,10:00测量的叶水势,在轻旱与对照下无显著差异,中、重旱下显著降低.叶水势能够直接反映作物的水分亏缺情况,叶水势对WUE的影响是通过影响气孔行为实现的,当干旱胁迫刚开始时叶片可通过气孔的部分关闭来减少水分丧失,从而使叶水势在一定范围内保持恒定(张文丽等,2006),因而叶水势在轻旱处理下与对照处理无明显差异.但叶水势发生轻微下降都将提高植物激素脱落酸(Abscisic Acid,ABA)浓度,引起叶片气孔的轻微关闭以防止蒸腾性水分流失(Laxa et al.,2019),即气孔导度降低在叶片水势还没有明显变化之前就已经发生,这种生存策略会减少叶片CO2的吸收量,最终导致净光合速率下降,从而轻旱处理下WUE小于对照处理.当植物遭受较强的干旱胁迫后,会优先考虑生存而非生产力,而角质层损失的水量决定其存活能力(Mukarram et al.,2021),气孔的关闭不仅为了防止植物失水过度,同时可以使叶片水分利用效率最大化(战吉宬等,2005).叶水势在中旱下显著减小,气孔进一步关闭,以水分损失最小化为主要目的使蒸腾速率急剧降低(赵福年等,2019),此时蒸腾对干旱的响应比光合反应更强烈,因此蒸腾速率下降幅度要快于光合速率,也正是光合作用和蒸腾速率间的耦合关系促使中旱胁迫下水分利用效率最高,即气孔导度对叶片在碳同化和水分耗散的调节中符合最优控制的原则,这是植物抗旱策略的重要组成部分.重旱胁迫会导致叶绿素功能障碍并破坏光合器官(Cai et al.,2020),气孔对叶水势失去调节能力,因而重旱下叶水势降到极低,WUE显著减小,这与蔡福等(2017)的研究一致. ...
Physiological limitations to photosynthetic carbon assimilation in cotton under water stress
1
2005
... 光合作用是地球生态系统一切生命活动的能量来源,也是作物产量形成的生理基础,其对环境变化十分敏感(卜令铎等,2010;李建查等,2018).作为植物最常见的生长逆境,干旱是抑制玉米生长和光合作用下降的重要原因(张仁和等,2011),限制着植物的光能利用率,甚至造成叶片结构的损伤(Mukarram et al.,2021),最终导致植物生物量及产量的下降(Jia et al.,2020).叶片光合作用对光的响应曲线是研究作物光合作用对环境变化响应的重要手段(李义博等,2017),可反映植物对不同光强的利用规律,并指示植物的生长情况及其抗逆性.通常以气孔导度、净光合速率、蒸腾速率及水分利用效率等指标来表征作物的光合特性(于文颖等,2015).气孔不仅是水分和光合原料进入细胞的关键通道,在干旱感知和信号传导过程中也发挥着不可或缺的作用(Laxa et al.,2019).Hetherington和Woodward(2003)研究指出,气孔面积仅占叶片面积的5%,但经由气孔蒸发的水分却高达70%.由此可见,气孔调节作为植物抵御胁迫环境的核心机制,对于维持植物体内水分平衡、保障光合作用正常进行意义重大.气孔导度作为衡量气孔变化特征的重要指标,其大小直接影响叶片中CO2的交换速率和水分的蒸腾速率(龚容和高琼,2015).适度水分亏缺下叶片可通过改变气孔的形态和气孔导度的大小来控制叶片内部CO2浓度和蒸腾速率,协调碳同化和水分消耗之间的关系(Ennahli and Earl,2005),同时影响叶片水分利用效率(Water Use Efficiency,WUE),最终形成高效的抗旱策略以适应干旱环境(李民青等,2023),随着干旱条件的进一步发展,作物生产可能出现环境约束向生理生态约束的转变,植物气孔可能关闭,导致叶绿体结构的变化和对植物膜系统的损害,因而阐明光合速率下降的生理机制对于准确识别作物干旱程度和制定对策至关重要(Song et al.,2020). ...
Stomatal conductance and photosynthesis
3
1982
... 植物光合作用主要受气孔和非气孔因素影响(Jia et al.,2020),前者指干旱胁迫导致的气孔导度降低,CO2进入叶片的通道受阻而使光合速率下降(Song et al.,2020),后者主要是叶肉细胞电子传递和光合磷酸化受阻,暗反应中相关酶活性降低,导致光合速率下降(Farquhar and Sharkey,1982).宋丰萍和蒙祖庆(2018)研究表明,随着干旱胁迫的逐步加剧、持续时间不断延长,气孔和非气孔因素往往会同时对植物光合作用产生影响,在这一过程中,非气孔因素最终会成为主导限制因子.基于此,麻雪艳和周广胜(2018)提出,气孔限制向非气孔限制的转折点,可作为判定玉米是否受到严重干旱胁迫的重要依据. ...
... 于5月30日春玉米七叶期开始时进行控水(即播种后第30天),随后开展各项观测工作.每天定时观测土壤水分、光合生理参数及叶水势等指标,直至植株萎蔫死亡,试验结束.在整个试验时段内,每日06:00和10:00采集第一片完全展开叶,使用WP4露点水势仪测定叶水势,每个处理设置3次重复(张健等,2019).在试验时段内,为确保测量数据的准确性与稳定性,选定每天上午09:00—11:00,利用Li-6400XT光合测量系统(LI-COR,Lincoln,USA)测定春玉米叶片的光合生理指标.选取玉米顶部第一片展开叶为测量对象,测量时指定使用人工红蓝光源,CO2浓度采用小钢瓶控制,样品室浓度设置为400 μmol·mol-1,叶室温度设置为25 ℃,在1 800 μmol·m-2·s-1的光强下充分诱导,待光合参数稳定后,利用自动测量程序进行测量,光合有效辐射(Photosynthetically Active Radiation,PAR)梯度设定为2 400、2 100、1 800、1 500、1 200、1 000、800、600、400、200、150、100、75、50、25、0 μmol·m-2·s-1,每个处理3次重复.测定内容包括:净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs),并计算气孔限制值(Ls)(Farquhar and Sharkey,1982)和瞬时水分利用效率(WUE),具体公式如下: ...
... 植物受到干旱胁迫时光合作用被限制,碳同化能力下降(Neto et al.,2017),光合速率的下降由气孔和非气孔因素共同决定(Jia et al.,2020).叶子飘等(2019)研究表明,当所有气孔开闭行为保持一致时,判断叶片光合速率下降是否由气孔因素主导,需依据胞间CO2浓度(Ci)和气孔限制值的变化趋势来判定,而非单纯关注Ci的下降幅度.根据Farquhar和Sharkey(1982)的观点,在Pn减小的情况下,Ci下降且Ls升高时,气孔限制占主导作用,当Ci升高且Ls下降时,非气孔因素占主导作用.本研究中,随着光照增强,轻旱胁迫下玉米Gs相较于对照处理显著下降,叶片Ci增加,Ls减小,根据Ci的变化方向判断此时以非气孔限制为主导因素使净光合速率降低.但许大全(2013)指出叶片气孔的不均关闭现象会使Ci恒定不变,水分胁迫条件下容易造成非气孔限制的假象,当作物突然遭受水分胁迫,植物为维持正常的叶水势,通过关闭部分气孔的方式减少水分蒸腾(Sharma and Zheng,2019),计算的Ci会高于或者等于实际值,进而造成非气孔限制的假象.Medrano等(2002)提出,可由光饱和的气孔导度作为干旱条件下光合作用的气孔和非气孔限制之间的拐点指标,最大气孔导度值在0.05 mol·m-2·s-1左右时即为拐点值,因而轻旱下主要是气孔限制影响了叶片碳同化能力,导致光合速率下降.中旱处理下玉米叶片Ci减小,Ls增加,Gs和Pn均显著下降,结合最大气孔导度拐点值,此时叶片净光合速率下降的因素以气孔限制为主导,非气孔限制逐渐增强.中旱处理下植物气孔的开放程度进一步降低,气孔阻力增加,限制了光合作用原料的供应,导致Ci降低(Song et al.,2020),最终使作物光合作用速率降低,叶片通过气孔调节的方式降低水分蒸腾,避免干旱对叶片结构的损伤,这可能是本文中旱处理下气孔限制始终占主导因素的原因,体现了植物适应胁迫环境的气孔调节机制,表明玉米属较耐旱的作物,在受到干旱胁迫后作物光合系统的自身调节能力较强,可忍耐一定程度的水分亏缺(陈图强等,2023).发生轻微干旱时,植物的抗氧化防御机制一般可以及时清除活性氧自由基以避免膜损伤(Jia et al.,2020),但重旱下土壤水分胁迫超出了植物可应对的阈值,叶片不能得到生存所需的最低水分,植物收支平衡被打破,叶肉细胞光合活性降低,抗氧化防御机理极其脆弱,活性氧的积累会破坏光系统(Sharma and Zheng,2019),植物无法维持正常生理活动.重旱下玉米叶片Ci显著增加,Ls明显减小,非气孔因素限制作物的光合作用,因而净光合速率等光合参数趋近于零. ...
The role of stomata in sensing and driving environmental change
1
2003
... 光合作用是地球生态系统一切生命活动的能量来源,也是作物产量形成的生理基础,其对环境变化十分敏感(卜令铎等,2010;李建查等,2018).作为植物最常见的生长逆境,干旱是抑制玉米生长和光合作用下降的重要原因(张仁和等,2011),限制着植物的光能利用率,甚至造成叶片结构的损伤(Mukarram et al.,2021),最终导致植物生物量及产量的下降(Jia et al.,2020).叶片光合作用对光的响应曲线是研究作物光合作用对环境变化响应的重要手段(李义博等,2017),可反映植物对不同光强的利用规律,并指示植物的生长情况及其抗逆性.通常以气孔导度、净光合速率、蒸腾速率及水分利用效率等指标来表征作物的光合特性(于文颖等,2015).气孔不仅是水分和光合原料进入细胞的关键通道,在干旱感知和信号传导过程中也发挥着不可或缺的作用(Laxa et al.,2019).Hetherington和Woodward(2003)研究指出,气孔面积仅占叶片面积的5%,但经由气孔蒸发的水分却高达70%.由此可见,气孔调节作为植物抵御胁迫环境的核心机制,对于维持植物体内水分平衡、保障光合作用正常进行意义重大.气孔导度作为衡量气孔变化特征的重要指标,其大小直接影响叶片中CO2的交换速率和水分的蒸腾速率(龚容和高琼,2015).适度水分亏缺下叶片可通过改变气孔的形态和气孔导度的大小来控制叶片内部CO2浓度和蒸腾速率,协调碳同化和水分消耗之间的关系(Ennahli and Earl,2005),同时影响叶片水分利用效率(Water Use Efficiency,WUE),最终形成高效的抗旱策略以适应干旱环境(李民青等,2023),随着干旱条件的进一步发展,作物生产可能出现环境约束向生理生态约束的转变,植物气孔可能关闭,导致叶绿体结构的变化和对植物膜系统的损害,因而阐明光合速率下降的生理机制对于准确识别作物干旱程度和制定对策至关重要(Song et al.,2020). ...
Response of photosynthetic performance to drought duration and re-watering in maize
4
2020
... 光合作用是地球生态系统一切生命活动的能量来源,也是作物产量形成的生理基础,其对环境变化十分敏感(卜令铎等,2010;李建查等,2018).作为植物最常见的生长逆境,干旱是抑制玉米生长和光合作用下降的重要原因(张仁和等,2011),限制着植物的光能利用率,甚至造成叶片结构的损伤(Mukarram et al.,2021),最终导致植物生物量及产量的下降(Jia et al.,2020).叶片光合作用对光的响应曲线是研究作物光合作用对环境变化响应的重要手段(李义博等,2017),可反映植物对不同光强的利用规律,并指示植物的生长情况及其抗逆性.通常以气孔导度、净光合速率、蒸腾速率及水分利用效率等指标来表征作物的光合特性(于文颖等,2015).气孔不仅是水分和光合原料进入细胞的关键通道,在干旱感知和信号传导过程中也发挥着不可或缺的作用(Laxa et al.,2019).Hetherington和Woodward(2003)研究指出,气孔面积仅占叶片面积的5%,但经由气孔蒸发的水分却高达70%.由此可见,气孔调节作为植物抵御胁迫环境的核心机制,对于维持植物体内水分平衡、保障光合作用正常进行意义重大.气孔导度作为衡量气孔变化特征的重要指标,其大小直接影响叶片中CO2的交换速率和水分的蒸腾速率(龚容和高琼,2015).适度水分亏缺下叶片可通过改变气孔的形态和气孔导度的大小来控制叶片内部CO2浓度和蒸腾速率,协调碳同化和水分消耗之间的关系(Ennahli and Earl,2005),同时影响叶片水分利用效率(Water Use Efficiency,WUE),最终形成高效的抗旱策略以适应干旱环境(李民青等,2023),随着干旱条件的进一步发展,作物生产可能出现环境约束向生理生态约束的转变,植物气孔可能关闭,导致叶绿体结构的变化和对植物膜系统的损害,因而阐明光合速率下降的生理机制对于准确识别作物干旱程度和制定对策至关重要(Song et al.,2020). ...
... 植物光合作用主要受气孔和非气孔因素影响(Jia et al.,2020),前者指干旱胁迫导致的气孔导度降低,CO2进入叶片的通道受阻而使光合速率下降(Song et al.,2020),后者主要是叶肉细胞电子传递和光合磷酸化受阻,暗反应中相关酶活性降低,导致光合速率下降(Farquhar and Sharkey,1982).宋丰萍和蒙祖庆(2018)研究表明,随着干旱胁迫的逐步加剧、持续时间不断延长,气孔和非气孔因素往往会同时对植物光合作用产生影响,在这一过程中,非气孔因素最终会成为主导限制因子.基于此,麻雪艳和周广胜(2018)提出,气孔限制向非气孔限制的转折点,可作为判定玉米是否受到严重干旱胁迫的重要依据. ...
... 植物受到干旱胁迫时光合作用被限制,碳同化能力下降(Neto et al.,2017),光合速率的下降由气孔和非气孔因素共同决定(Jia et al.,2020).叶子飘等(2019)研究表明,当所有气孔开闭行为保持一致时,判断叶片光合速率下降是否由气孔因素主导,需依据胞间CO2浓度(Ci)和气孔限制值的变化趋势来判定,而非单纯关注Ci的下降幅度.根据Farquhar和Sharkey(1982)的观点,在Pn减小的情况下,Ci下降且Ls升高时,气孔限制占主导作用,当Ci升高且Ls下降时,非气孔因素占主导作用.本研究中,随着光照增强,轻旱胁迫下玉米Gs相较于对照处理显著下降,叶片Ci增加,Ls减小,根据Ci的变化方向判断此时以非气孔限制为主导因素使净光合速率降低.但许大全(2013)指出叶片气孔的不均关闭现象会使Ci恒定不变,水分胁迫条件下容易造成非气孔限制的假象,当作物突然遭受水分胁迫,植物为维持正常的叶水势,通过关闭部分气孔的方式减少水分蒸腾(Sharma and Zheng,2019),计算的Ci会高于或者等于实际值,进而造成非气孔限制的假象.Medrano等(2002)提出,可由光饱和的气孔导度作为干旱条件下光合作用的气孔和非气孔限制之间的拐点指标,最大气孔导度值在0.05 mol·m-2·s-1左右时即为拐点值,因而轻旱下主要是气孔限制影响了叶片碳同化能力,导致光合速率下降.中旱处理下玉米叶片Ci减小,Ls增加,Gs和Pn均显著下降,结合最大气孔导度拐点值,此时叶片净光合速率下降的因素以气孔限制为主导,非气孔限制逐渐增强.中旱处理下植物气孔的开放程度进一步降低,气孔阻力增加,限制了光合作用原料的供应,导致Ci降低(Song et al.,2020),最终使作物光合作用速率降低,叶片通过气孔调节的方式降低水分蒸腾,避免干旱对叶片结构的损伤,这可能是本文中旱处理下气孔限制始终占主导因素的原因,体现了植物适应胁迫环境的气孔调节机制,表明玉米属较耐旱的作物,在受到干旱胁迫后作物光合系统的自身调节能力较强,可忍耐一定程度的水分亏缺(陈图强等,2023).发生轻微干旱时,植物的抗氧化防御机制一般可以及时清除活性氧自由基以避免膜损伤(Jia et al.,2020),但重旱下土壤水分胁迫超出了植物可应对的阈值,叶片不能得到生存所需的最低水分,植物收支平衡被打破,叶肉细胞光合活性降低,抗氧化防御机理极其脆弱,活性氧的积累会破坏光系统(Sharma and Zheng,2019),植物无法维持正常生理活动.重旱下玉米叶片Ci显著增加,Ls明显减小,非气孔因素限制作物的光合作用,因而净光合速率等光合参数趋近于零. ...
... ).发生轻微干旱时,植物的抗氧化防御机制一般可以及时清除活性氧自由基以避免膜损伤(Jia et al.,2020),但重旱下土壤水分胁迫超出了植物可应对的阈值,叶片不能得到生存所需的最低水分,植物收支平衡被打破,叶肉细胞光合活性降低,抗氧化防御机理极其脆弱,活性氧的积累会破坏光系统(Sharma and Zheng,2019),植物无法维持正常生理活动.重旱下玉米叶片Ci显著增加,Ls明显减小,非气孔因素限制作物的光合作用,因而净光合速率等光合参数趋近于零. ...
The role of the plant antioxidant system in drought tolerance
2
2019
... 光合作用是地球生态系统一切生命活动的能量来源,也是作物产量形成的生理基础,其对环境变化十分敏感(卜令铎等,2010;李建查等,2018).作为植物最常见的生长逆境,干旱是抑制玉米生长和光合作用下降的重要原因(张仁和等,2011),限制着植物的光能利用率,甚至造成叶片结构的损伤(Mukarram et al.,2021),最终导致植物生物量及产量的下降(Jia et al.,2020).叶片光合作用对光的响应曲线是研究作物光合作用对环境变化响应的重要手段(李义博等,2017),可反映植物对不同光强的利用规律,并指示植物的生长情况及其抗逆性.通常以气孔导度、净光合速率、蒸腾速率及水分利用效率等指标来表征作物的光合特性(于文颖等,2015).气孔不仅是水分和光合原料进入细胞的关键通道,在干旱感知和信号传导过程中也发挥着不可或缺的作用(Laxa et al.,2019).Hetherington和Woodward(2003)研究指出,气孔面积仅占叶片面积的5%,但经由气孔蒸发的水分却高达70%.由此可见,气孔调节作为植物抵御胁迫环境的核心机制,对于维持植物体内水分平衡、保障光合作用正常进行意义重大.气孔导度作为衡量气孔变化特征的重要指标,其大小直接影响叶片中CO2的交换速率和水分的蒸腾速率(龚容和高琼,2015).适度水分亏缺下叶片可通过改变气孔的形态和气孔导度的大小来控制叶片内部CO2浓度和蒸腾速率,协调碳同化和水分消耗之间的关系(Ennahli and Earl,2005),同时影响叶片水分利用效率(Water Use Efficiency,WUE),最终形成高效的抗旱策略以适应干旱环境(李民青等,2023),随着干旱条件的进一步发展,作物生产可能出现环境约束向生理生态约束的转变,植物气孔可能关闭,导致叶绿体结构的变化和对植物膜系统的损害,因而阐明光合速率下降的生理机制对于准确识别作物干旱程度和制定对策至关重要(Song et al.,2020). ...
... 水分利用效率(WUE)是植物基本的生理生态特征,高效的WUE有利于维持植物的碳水平衡和适应干旱环境(李民青等,2023).叶片尺度的WUE由光合作用与蒸腾作用共同决定,因而影响作物这两个生理过程的因素均会引起叶片WUE的变化(姜寒冰等,2019).本研究中,随着干旱胁迫的增加,玉米叶片的Pn和Tr均逐步降低,WUE最终表现为中旱与对照无显著差异,轻旱和重旱显著降低,10:00测量的叶水势,在轻旱与对照下无显著差异,中、重旱下显著降低.叶水势能够直接反映作物的水分亏缺情况,叶水势对WUE的影响是通过影响气孔行为实现的,当干旱胁迫刚开始时叶片可通过气孔的部分关闭来减少水分丧失,从而使叶水势在一定范围内保持恒定(张文丽等,2006),因而叶水势在轻旱处理下与对照处理无明显差异.但叶水势发生轻微下降都将提高植物激素脱落酸(Abscisic Acid,ABA)浓度,引起叶片气孔的轻微关闭以防止蒸腾性水分流失(Laxa et al.,2019),即气孔导度降低在叶片水势还没有明显变化之前就已经发生,这种生存策略会减少叶片CO2的吸收量,最终导致净光合速率下降,从而轻旱处理下WUE小于对照处理.当植物遭受较强的干旱胁迫后,会优先考虑生存而非生产力,而角质层损失的水量决定其存活能力(Mukarram et al.,2021),气孔的关闭不仅为了防止植物失水过度,同时可以使叶片水分利用效率最大化(战吉宬等,2005).叶水势在中旱下显著减小,气孔进一步关闭,以水分损失最小化为主要目的使蒸腾速率急剧降低(赵福年等,2019),此时蒸腾对干旱的响应比光合反应更强烈,因此蒸腾速率下降幅度要快于光合速率,也正是光合作用和蒸腾速率间的耦合关系促使中旱胁迫下水分利用效率最高,即气孔导度对叶片在碳同化和水分耗散的调节中符合最优控制的原则,这是植物抗旱策略的重要组成部分.重旱胁迫会导致叶绿素功能障碍并破坏光合器官(Cai et al.,2020),气孔对叶水势失去调节能力,因而重旱下叶水势降到极低,WUE显著减小,这与蔡福等(2017)的研究一致. ...
Greater sensitivity to drought accompanies maize yield increase in the U.S. Midwest
1
2014
... 干旱是农作物生长的主要制约因素之一(Lobell et al.,2014),玉米作为中国种植范围最广的谷类作物,在农业及国民经济中占据着重要地位(宋贺等,2019).全球气候变化背景下水热格局的改变,加剧了干旱等非生物胁迫因子对作物生长和产量的限制作用(李建查等,2018).近年来,光照、水分胁迫等环境因子对作物光合生理特征的影响得到了广泛关注(张仁和等,2011;Sharma and Zheng,2019;姜鹏等,2023),不同强度干旱过程对玉米生长发育、光合生理的影响也有较多研究,主要集中在拔节期和抽雄期(于文颖等,2015;宋贺等,2019;杨阳等,2022).萌发期和苗期(前期干旱)是春玉米干旱发生频率最高的阶段(任宗悦等,2020),特别是玉米营养生长阶段的七叶期(齐月等,2023),此时遭遇干旱会严重抑制春玉米的生长发育(卜令铎等,2010;Song et al.,2019).然而,目前针对干旱胁迫下春玉米七叶期的光合响应特征及其影响机制的研究尚显不足,因此本文通过研究春玉米七叶期光合参数变化特征及其对干旱的响应,试图明晰不同干旱程度下春玉米七叶期的叶片生理抗旱机制,以期为农业生产过程中春玉米生长前期的水分管理提供理论依据. ...
Regulation of photosynthesis of C3 plants in response to progressive drought: Stomatal conductance as a reference parameter
1
2002
... 植物受到干旱胁迫时光合作用被限制,碳同化能力下降(Neto et al.,2017),光合速率的下降由气孔和非气孔因素共同决定(Jia et al.,2020).叶子飘等(2019)研究表明,当所有气孔开闭行为保持一致时,判断叶片光合速率下降是否由气孔因素主导,需依据胞间CO2浓度(Ci)和气孔限制值的变化趋势来判定,而非单纯关注Ci的下降幅度.根据Farquhar和Sharkey(1982)的观点,在Pn减小的情况下,Ci下降且Ls升高时,气孔限制占主导作用,当Ci升高且Ls下降时,非气孔因素占主导作用.本研究中,随着光照增强,轻旱胁迫下玉米Gs相较于对照处理显著下降,叶片Ci增加,Ls减小,根据Ci的变化方向判断此时以非气孔限制为主导因素使净光合速率降低.但许大全(2013)指出叶片气孔的不均关闭现象会使Ci恒定不变,水分胁迫条件下容易造成非气孔限制的假象,当作物突然遭受水分胁迫,植物为维持正常的叶水势,通过关闭部分气孔的方式减少水分蒸腾(Sharma and Zheng,2019),计算的Ci会高于或者等于实际值,进而造成非气孔限制的假象.Medrano等(2002)提出,可由光饱和的气孔导度作为干旱条件下光合作用的气孔和非气孔限制之间的拐点指标,最大气孔导度值在0.05 mol·m-2·s-1左右时即为拐点值,因而轻旱下主要是气孔限制影响了叶片碳同化能力,导致光合速率下降.中旱处理下玉米叶片Ci减小,Ls增加,Gs和Pn均显著下降,结合最大气孔导度拐点值,此时叶片净光合速率下降的因素以气孔限制为主导,非气孔限制逐渐增强.中旱处理下植物气孔的开放程度进一步降低,气孔阻力增加,限制了光合作用原料的供应,导致Ci降低(Song et al.,2020),最终使作物光合作用速率降低,叶片通过气孔调节的方式降低水分蒸腾,避免干旱对叶片结构的损伤,这可能是本文中旱处理下气孔限制始终占主导因素的原因,体现了植物适应胁迫环境的气孔调节机制,表明玉米属较耐旱的作物,在受到干旱胁迫后作物光合系统的自身调节能力较强,可忍耐一定程度的水分亏缺(陈图强等,2023).发生轻微干旱时,植物的抗氧化防御机制一般可以及时清除活性氧自由基以避免膜损伤(Jia et al.,2020),但重旱下土壤水分胁迫超出了植物可应对的阈值,叶片不能得到生存所需的最低水分,植物收支平衡被打破,叶肉细胞光合活性降低,抗氧化防御机理极其脆弱,活性氧的积累会破坏光系统(Sharma and Zheng,2019),植物无法维持正常生理活动.重旱下玉米叶片Ci显著增加,Ls明显减小,非气孔因素限制作物的光合作用,因而净光合速率等光合参数趋近于零. ...
Drought: Sensing, signalling, effects and tolerance in higher plants
2
2021
... 光合作用是地球生态系统一切生命活动的能量来源,也是作物产量形成的生理基础,其对环境变化十分敏感(卜令铎等,2010;李建查等,2018).作为植物最常见的生长逆境,干旱是抑制玉米生长和光合作用下降的重要原因(张仁和等,2011),限制着植物的光能利用率,甚至造成叶片结构的损伤(Mukarram et al.,2021),最终导致植物生物量及产量的下降(Jia et al.,2020).叶片光合作用对光的响应曲线是研究作物光合作用对环境变化响应的重要手段(李义博等,2017),可反映植物对不同光强的利用规律,并指示植物的生长情况及其抗逆性.通常以气孔导度、净光合速率、蒸腾速率及水分利用效率等指标来表征作物的光合特性(于文颖等,2015).气孔不仅是水分和光合原料进入细胞的关键通道,在干旱感知和信号传导过程中也发挥着不可或缺的作用(Laxa et al.,2019).Hetherington和Woodward(2003)研究指出,气孔面积仅占叶片面积的5%,但经由气孔蒸发的水分却高达70%.由此可见,气孔调节作为植物抵御胁迫环境的核心机制,对于维持植物体内水分平衡、保障光合作用正常进行意义重大.气孔导度作为衡量气孔变化特征的重要指标,其大小直接影响叶片中CO2的交换速率和水分的蒸腾速率(龚容和高琼,2015).适度水分亏缺下叶片可通过改变气孔的形态和气孔导度的大小来控制叶片内部CO2浓度和蒸腾速率,协调碳同化和水分消耗之间的关系(Ennahli and Earl,2005),同时影响叶片水分利用效率(Water Use Efficiency,WUE),最终形成高效的抗旱策略以适应干旱环境(李民青等,2023),随着干旱条件的进一步发展,作物生产可能出现环境约束向生理生态约束的转变,植物气孔可能关闭,导致叶绿体结构的变化和对植物膜系统的损害,因而阐明光合速率下降的生理机制对于准确识别作物干旱程度和制定对策至关重要(Song et al.,2020). ...
... 水分利用效率(WUE)是植物基本的生理生态特征,高效的WUE有利于维持植物的碳水平衡和适应干旱环境(李民青等,2023).叶片尺度的WUE由光合作用与蒸腾作用共同决定,因而影响作物这两个生理过程的因素均会引起叶片WUE的变化(姜寒冰等,2019).本研究中,随着干旱胁迫的增加,玉米叶片的Pn和Tr均逐步降低,WUE最终表现为中旱与对照无显著差异,轻旱和重旱显著降低,10:00测量的叶水势,在轻旱与对照下无显著差异,中、重旱下显著降低.叶水势能够直接反映作物的水分亏缺情况,叶水势对WUE的影响是通过影响气孔行为实现的,当干旱胁迫刚开始时叶片可通过气孔的部分关闭来减少水分丧失,从而使叶水势在一定范围内保持恒定(张文丽等,2006),因而叶水势在轻旱处理下与对照处理无明显差异.但叶水势发生轻微下降都将提高植物激素脱落酸(Abscisic Acid,ABA)浓度,引起叶片气孔的轻微关闭以防止蒸腾性水分流失(Laxa et al.,2019),即气孔导度降低在叶片水势还没有明显变化之前就已经发生,这种生存策略会减少叶片CO2的吸收量,最终导致净光合速率下降,从而轻旱处理下WUE小于对照处理.当植物遭受较强的干旱胁迫后,会优先考虑生存而非生产力,而角质层损失的水量决定其存活能力(Mukarram et al.,2021),气孔的关闭不仅为了防止植物失水过度,同时可以使叶片水分利用效率最大化(战吉宬等,2005).叶水势在中旱下显著减小,气孔进一步关闭,以水分损失最小化为主要目的使蒸腾速率急剧降低(赵福年等,2019),此时蒸腾对干旱的响应比光合反应更强烈,因此蒸腾速率下降幅度要快于光合速率,也正是光合作用和蒸腾速率间的耦合关系促使中旱胁迫下水分利用效率最高,即气孔导度对叶片在碳同化和水分耗散的调节中符合最优控制的原则,这是植物抗旱策略的重要组成部分.重旱胁迫会导致叶绿素功能障碍并破坏光合器官(Cai et al.,2020),气孔对叶水势失去调节能力,因而重旱下叶水势降到极低,WUE显著减小,这与蔡福等(2017)的研究一致. ...
Cyclic electron flow, NPQ and photorespiration are crucial for the establishment of young plants of Ricinus communis and Jatropha curcas exposed to drought
1
2017
... 植物受到干旱胁迫时光合作用被限制,碳同化能力下降(Neto et al.,2017),光合速率的下降由气孔和非气孔因素共同决定(Jia et al.,2020).叶子飘等(2019)研究表明,当所有气孔开闭行为保持一致时,判断叶片光合速率下降是否由气孔因素主导,需依据胞间CO2浓度(Ci)和气孔限制值的变化趋势来判定,而非单纯关注Ci的下降幅度.根据Farquhar和Sharkey(1982)的观点,在Pn减小的情况下,Ci下降且Ls升高时,气孔限制占主导作用,当Ci升高且Ls下降时,非气孔因素占主导作用.本研究中,随着光照增强,轻旱胁迫下玉米Gs相较于对照处理显著下降,叶片Ci增加,Ls减小,根据Ci的变化方向判断此时以非气孔限制为主导因素使净光合速率降低.但许大全(2013)指出叶片气孔的不均关闭现象会使Ci恒定不变,水分胁迫条件下容易造成非气孔限制的假象,当作物突然遭受水分胁迫,植物为维持正常的叶水势,通过关闭部分气孔的方式减少水分蒸腾(Sharma and Zheng,2019),计算的Ci会高于或者等于实际值,进而造成非气孔限制的假象.Medrano等(2002)提出,可由光饱和的气孔导度作为干旱条件下光合作用的气孔和非气孔限制之间的拐点指标,最大气孔导度值在0.05 mol·m-2·s-1左右时即为拐点值,因而轻旱下主要是气孔限制影响了叶片碳同化能力,导致光合速率下降.中旱处理下玉米叶片Ci减小,Ls增加,Gs和Pn均显著下降,结合最大气孔导度拐点值,此时叶片净光合速率下降的因素以气孔限制为主导,非气孔限制逐渐增强.中旱处理下植物气孔的开放程度进一步降低,气孔阻力增加,限制了光合作用原料的供应,导致Ci降低(Song et al.,2020),最终使作物光合作用速率降低,叶片通过气孔调节的方式降低水分蒸腾,避免干旱对叶片结构的损伤,这可能是本文中旱处理下气孔限制始终占主导因素的原因,体现了植物适应胁迫环境的气孔调节机制,表明玉米属较耐旱的作物,在受到干旱胁迫后作物光合系统的自身调节能力较强,可忍耐一定程度的水分亏缺(陈图强等,2023).发生轻微干旱时,植物的抗氧化防御机制一般可以及时清除活性氧自由基以避免膜损伤(Jia et al.,2020),但重旱下土壤水分胁迫超出了植物可应对的阈值,叶片不能得到生存所需的最低水分,植物收支平衡被打破,叶肉细胞光合活性降低,抗氧化防御机理极其脆弱,活性氧的积累会破坏光系统(Sharma and Zheng,2019),植物无法维持正常生理活动.重旱下玉米叶片Ci显著增加,Ls明显减小,非气孔因素限制作物的光合作用,因而净光合速率等光合参数趋近于零. ...
Melatonin mediated regulation of drought stress: Physiological and molecular aspects
3
2019
... 干旱是农作物生长的主要制约因素之一(Lobell et al.,2014),玉米作为中国种植范围最广的谷类作物,在农业及国民经济中占据着重要地位(宋贺等,2019).全球气候变化背景下水热格局的改变,加剧了干旱等非生物胁迫因子对作物生长和产量的限制作用(李建查等,2018).近年来,光照、水分胁迫等环境因子对作物光合生理特征的影响得到了广泛关注(张仁和等,2011;Sharma and Zheng,2019;姜鹏等,2023),不同强度干旱过程对玉米生长发育、光合生理的影响也有较多研究,主要集中在拔节期和抽雄期(于文颖等,2015;宋贺等,2019;杨阳等,2022).萌发期和苗期(前期干旱)是春玉米干旱发生频率最高的阶段(任宗悦等,2020),特别是玉米营养生长阶段的七叶期(齐月等,2023),此时遭遇干旱会严重抑制春玉米的生长发育(卜令铎等,2010;Song et al.,2019).然而,目前针对干旱胁迫下春玉米七叶期的光合响应特征及其影响机制的研究尚显不足,因此本文通过研究春玉米七叶期光合参数变化特征及其对干旱的响应,试图明晰不同干旱程度下春玉米七叶期的叶片生理抗旱机制,以期为农业生产过程中春玉米生长前期的水分管理提供理论依据. ...
... 植物受到干旱胁迫时光合作用被限制,碳同化能力下降(Neto et al.,2017),光合速率的下降由气孔和非气孔因素共同决定(Jia et al.,2020).叶子飘等(2019)研究表明,当所有气孔开闭行为保持一致时,判断叶片光合速率下降是否由气孔因素主导,需依据胞间CO2浓度(Ci)和气孔限制值的变化趋势来判定,而非单纯关注Ci的下降幅度.根据Farquhar和Sharkey(1982)的观点,在Pn减小的情况下,Ci下降且Ls升高时,气孔限制占主导作用,当Ci升高且Ls下降时,非气孔因素占主导作用.本研究中,随着光照增强,轻旱胁迫下玉米Gs相较于对照处理显著下降,叶片Ci增加,Ls减小,根据Ci的变化方向判断此时以非气孔限制为主导因素使净光合速率降低.但许大全(2013)指出叶片气孔的不均关闭现象会使Ci恒定不变,水分胁迫条件下容易造成非气孔限制的假象,当作物突然遭受水分胁迫,植物为维持正常的叶水势,通过关闭部分气孔的方式减少水分蒸腾(Sharma and Zheng,2019),计算的Ci会高于或者等于实际值,进而造成非气孔限制的假象.Medrano等(2002)提出,可由光饱和的气孔导度作为干旱条件下光合作用的气孔和非气孔限制之间的拐点指标,最大气孔导度值在0.05 mol·m-2·s-1左右时即为拐点值,因而轻旱下主要是气孔限制影响了叶片碳同化能力,导致光合速率下降.中旱处理下玉米叶片Ci减小,Ls增加,Gs和Pn均显著下降,结合最大气孔导度拐点值,此时叶片净光合速率下降的因素以气孔限制为主导,非气孔限制逐渐增强.中旱处理下植物气孔的开放程度进一步降低,气孔阻力增加,限制了光合作用原料的供应,导致Ci降低(Song et al.,2020),最终使作物光合作用速率降低,叶片通过气孔调节的方式降低水分蒸腾,避免干旱对叶片结构的损伤,这可能是本文中旱处理下气孔限制始终占主导因素的原因,体现了植物适应胁迫环境的气孔调节机制,表明玉米属较耐旱的作物,在受到干旱胁迫后作物光合系统的自身调节能力较强,可忍耐一定程度的水分亏缺(陈图强等,2023).发生轻微干旱时,植物的抗氧化防御机制一般可以及时清除活性氧自由基以避免膜损伤(Jia et al.,2020),但重旱下土壤水分胁迫超出了植物可应对的阈值,叶片不能得到生存所需的最低水分,植物收支平衡被打破,叶肉细胞光合活性降低,抗氧化防御机理极其脆弱,活性氧的积累会破坏光系统(Sharma and Zheng,2019),植物无法维持正常生理活动.重旱下玉米叶片Ci显著增加,Ls明显减小,非气孔因素限制作物的光合作用,因而净光合速率等光合参数趋近于零. ...
... ),但重旱下土壤水分胁迫超出了植物可应对的阈值,叶片不能得到生存所需的最低水分,植物收支平衡被打破,叶肉细胞光合活性降低,抗氧化防御机理极其脆弱,活性氧的积累会破坏光系统(Sharma and Zheng,2019),植物无法维持正常生理活动.重旱下玉米叶片Ci显著增加,Ls明显减小,非气孔因素限制作物的光合作用,因而净光合速率等光合参数趋近于零. ...
Effects of severe water stress on maize growth processes in the field
1
2019
... 干旱是农作物生长的主要制约因素之一(Lobell et al.,2014),玉米作为中国种植范围最广的谷类作物,在农业及国民经济中占据着重要地位(宋贺等,2019).全球气候变化背景下水热格局的改变,加剧了干旱等非生物胁迫因子对作物生长和产量的限制作用(李建查等,2018).近年来,光照、水分胁迫等环境因子对作物光合生理特征的影响得到了广泛关注(张仁和等,2011;Sharma and Zheng,2019;姜鹏等,2023),不同强度干旱过程对玉米生长发育、光合生理的影响也有较多研究,主要集中在拔节期和抽雄期(于文颖等,2015;宋贺等,2019;杨阳等,2022).萌发期和苗期(前期干旱)是春玉米干旱发生频率最高的阶段(任宗悦等,2020),特别是玉米营养生长阶段的七叶期(齐月等,2023),此时遭遇干旱会严重抑制春玉米的生长发育(卜令铎等,2010;Song et al.,2019).然而,目前针对干旱胁迫下春玉米七叶期的光合响应特征及其影响机制的研究尚显不足,因此本文通过研究春玉米七叶期光合参数变化特征及其对干旱的响应,试图明晰不同干旱程度下春玉米七叶期的叶片生理抗旱机制,以期为农业生产过程中春玉米生长前期的水分管理提供理论依据. ...
Stomatal limitations to photosynthesis and their critical water conditions in different growth stages of maize under water stress
3
2020
... 光合作用是地球生态系统一切生命活动的能量来源,也是作物产量形成的生理基础,其对环境变化十分敏感(卜令铎等,2010;李建查等,2018).作为植物最常见的生长逆境,干旱是抑制玉米生长和光合作用下降的重要原因(张仁和等,2011),限制着植物的光能利用率,甚至造成叶片结构的损伤(Mukarram et al.,2021),最终导致植物生物量及产量的下降(Jia et al.,2020).叶片光合作用对光的响应曲线是研究作物光合作用对环境变化响应的重要手段(李义博等,2017),可反映植物对不同光强的利用规律,并指示植物的生长情况及其抗逆性.通常以气孔导度、净光合速率、蒸腾速率及水分利用效率等指标来表征作物的光合特性(于文颖等,2015).气孔不仅是水分和光合原料进入细胞的关键通道,在干旱感知和信号传导过程中也发挥着不可或缺的作用(Laxa et al.,2019).Hetherington和Woodward(2003)研究指出,气孔面积仅占叶片面积的5%,但经由气孔蒸发的水分却高达70%.由此可见,气孔调节作为植物抵御胁迫环境的核心机制,对于维持植物体内水分平衡、保障光合作用正常进行意义重大.气孔导度作为衡量气孔变化特征的重要指标,其大小直接影响叶片中CO2的交换速率和水分的蒸腾速率(龚容和高琼,2015).适度水分亏缺下叶片可通过改变气孔的形态和气孔导度的大小来控制叶片内部CO2浓度和蒸腾速率,协调碳同化和水分消耗之间的关系(Ennahli and Earl,2005),同时影响叶片水分利用效率(Water Use Efficiency,WUE),最终形成高效的抗旱策略以适应干旱环境(李民青等,2023),随着干旱条件的进一步发展,作物生产可能出现环境约束向生理生态约束的转变,植物气孔可能关闭,导致叶绿体结构的变化和对植物膜系统的损害,因而阐明光合速率下降的生理机制对于准确识别作物干旱程度和制定对策至关重要(Song et al.,2020). ...
... 植物光合作用主要受气孔和非气孔因素影响(Jia et al.,2020),前者指干旱胁迫导致的气孔导度降低,CO2进入叶片的通道受阻而使光合速率下降(Song et al.,2020),后者主要是叶肉细胞电子传递和光合磷酸化受阻,暗反应中相关酶活性降低,导致光合速率下降(Farquhar and Sharkey,1982).宋丰萍和蒙祖庆(2018)研究表明,随着干旱胁迫的逐步加剧、持续时间不断延长,气孔和非气孔因素往往会同时对植物光合作用产生影响,在这一过程中,非气孔因素最终会成为主导限制因子.基于此,麻雪艳和周广胜(2018)提出,气孔限制向非气孔限制的转折点,可作为判定玉米是否受到严重干旱胁迫的重要依据. ...
... 植物受到干旱胁迫时光合作用被限制,碳同化能力下降(Neto et al.,2017),光合速率的下降由气孔和非气孔因素共同决定(Jia et al.,2020).叶子飘等(2019)研究表明,当所有气孔开闭行为保持一致时,判断叶片光合速率下降是否由气孔因素主导,需依据胞间CO2浓度(Ci)和气孔限制值的变化趋势来判定,而非单纯关注Ci的下降幅度.根据Farquhar和Sharkey(1982)的观点,在Pn减小的情况下,Ci下降且Ls升高时,气孔限制占主导作用,当Ci升高且Ls下降时,非气孔因素占主导作用.本研究中,随着光照增强,轻旱胁迫下玉米Gs相较于对照处理显著下降,叶片Ci增加,Ls减小,根据Ci的变化方向判断此时以非气孔限制为主导因素使净光合速率降低.但许大全(2013)指出叶片气孔的不均关闭现象会使Ci恒定不变,水分胁迫条件下容易造成非气孔限制的假象,当作物突然遭受水分胁迫,植物为维持正常的叶水势,通过关闭部分气孔的方式减少水分蒸腾(Sharma and Zheng,2019),计算的Ci会高于或者等于实际值,进而造成非气孔限制的假象.Medrano等(2002)提出,可由光饱和的气孔导度作为干旱条件下光合作用的气孔和非气孔限制之间的拐点指标,最大气孔导度值在0.05 mol·m-2·s-1左右时即为拐点值,因而轻旱下主要是气孔限制影响了叶片碳同化能力,导致光合速率下降.中旱处理下玉米叶片Ci减小,Ls增加,Gs和Pn均显著下降,结合最大气孔导度拐点值,此时叶片净光合速率下降的因素以气孔限制为主导,非气孔限制逐渐增强.中旱处理下植物气孔的开放程度进一步降低,气孔阻力增加,限制了光合作用原料的供应,导致Ci降低(Song et al.,2020),最终使作物光合作用速率降低,叶片通过气孔调节的方式降低水分蒸腾,避免干旱对叶片结构的损伤,这可能是本文中旱处理下气孔限制始终占主导因素的原因,体现了植物适应胁迫环境的气孔调节机制,表明玉米属较耐旱的作物,在受到干旱胁迫后作物光合系统的自身调节能力较强,可忍耐一定程度的水分亏缺(陈图强等,2023).发生轻微干旱时,植物的抗氧化防御机制一般可以及时清除活性氧自由基以避免膜损伤(Jia et al.,2020),但重旱下土壤水分胁迫超出了植物可应对的阈值,叶片不能得到生存所需的最低水分,植物收支平衡被打破,叶肉细胞光合活性降低,抗氧化防御机理极其脆弱,活性氧的积累会破坏光系统(Sharma and Zheng,2019),植物无法维持正常生理活动.重旱下玉米叶片Ci显著增加,Ls明显减小,非气孔因素限制作物的光合作用,因而净光合速率等光合参数趋近于零. ...
Quantifying light response of leaf-scale water use efficiency and its interrelationships with photosynthesis and stomatal conductance in C3 and C4 species





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}