

0 引言
气候变化会对自然生态和社会经济系统产生深远影响,是当前全球最关注的问题之一。农业生态系统是最易受气候变化影响的生态系统之一(Meza and Silva,2009;薛思嘉等,2021)。有研究表明,气候变化对粮食产量的不利影响明显高于有利影响,如气候变化导致的小麦和玉米减产率分别为1.9%·(10 a)-1、1.2%·(10 a)-1(秦大河,2014)。联合国粮农组织预测,到2050年全球人口将达到91亿,粮食产量需在现有基础上至少增加70%才能满足需求(王丕屹,2009)。粮食减产与人口不断增长相叠加,使得粮食安全面临更大的挑战。观测数据表明,气候变化以平均气温显著上升为主要特征,而温度是作物生长发育过程中最敏感的气候因子,作物通过调整形态特征(如株高、叶面积、叶干重、根系形态及生物量等)来适应温度变化,这些特征指标不仅反映了植物与环境的关系,也是植物适应环境及获取资源能力的衡量(孟婷婷等,2007;刘晓娟和马克平,2015)。
冬小麦是关中平原的主要粮食作物之一,属喜凉作物,其生育期经历秋、冬、春三季,多处于较低温度环境中,因此对气候变暖有明显的生态响应。如增温对不同区域冬小麦株高的影响具有差异性(耿婷等,2014):在增温条件下,徐州冬小麦株高高于对照组,而丹阳冬小麦则低于对照组;增温可能对冬小麦株高有促进作用(房世波等,2010;田云录等,2011),也可能抑制株高(曹嘉晨等,2018)。夜间增温可以提高叶面积(杨锦浩等,2022),并随着增温强度增加,冬小麦叶面积明显增大(万敬敬,2017)。增温通常使物候期提前,但存在时空差异(张宸赫等,2016;郑孟静等,2022),He等(2015)研究表明温度升高使中国冬小麦的越冬期、开花期和成熟期提前,加速其生长发育过程。在一定生育阶段内,增温缩短了冬小麦越冬—拔节的生育进程,但延长了灌浆期的持续时间(谭凯炎等,2017;郑孟静等,2022),从而可能提高小麦产量(童思思等,2023)。增温不仅改变植物地上功能性状,还会促使根系形态发生相应调整,以提升养分获取能力(许璐璐等,2020)。一般认为增温在适宜范围内可促进小麦根体积、根表面积和根长的增加(张彬,2010),但当温度超过最适范围时,增温会抑制根系生长(Drennan and Nobel,1998),表现为根系生物量、总根长和代谢活性下降(Huang et al.,2012),增温对作物根系生长的影响因品种和耕作制度而异(Hou et al.,2018)。在干物质积累方面,生育中期(拔节—孕穗期)后,温度升高明显抑制春小麦各器官及总生物量的增加(张凯等,2016)、降低水稻各器官的干物质积累量(李春华等,2016);而拔节期前的增温处理可提高小麦成熟期各器官的干物质分配量(杨锦浩等,2022)。总之,增温对小麦地上、地下农艺性状的影响不同,最终影响干物质的转运方式。对于关中平原这一小麦主产区,模拟增温条件的研究需重点关注增温幅度对小麦农艺性状的具体影响,特别是营养生长期和生殖生长期增温对小麦的促进或抑制作用。此外,增温是否有利于穗干物质的增加仍需进一步研究,因为穗干物质的运转直接决定小麦产量,而产量下降将对粮食安全构成威胁(IPCC,2023)。
在气候变暖背景下,研究关中地区小麦的生长发育响应特征是亟待解决的科学问题。本研究通过模拟增温条件观测小麦生长过程,旨在探索气候变暖对小麦生长发育的影响,以期为未来气候条件下关中地区的粮食安全评估提供重要依据,并为制定应对气候变化、保障粮食安全的生产策略提供科学支撑。
1 数据与方法
1.1 试验设计
1.1.1 试验场概况
试验在秦岭-黄土高原过渡带的水土要素耦合与生物资源保育观测研究站进行。该观测站位于陕西省渭南市潼关县秦东镇小寨子村(110.25°E,34.59°N),海拔510 m,属温带大陆性气候,夏季湿热、冬季干冷,多年平均气温为14.7 ℃,多年平均降水量为601.9 mm,降水主要集中在7—9月(图1)。土壤类型为塿土,以黄土为母质,土质细腻均匀,肥力中等。0~30 cm土层有机质、全氮、总磷、总钾的含量分别为12.40、0.75、1.01、1.76 g·kg-1,速效磷、速效钾分别为6.7、151.0 mg·kg-1,pH值为7.98。小麦试验场分为两块,一块设置增温装置开顶式气室(Open Top Chamber,OTC),设有6个平行样;另一块为对照试验区(Control Check,CK),设有3个平行样。两组处理组均采用相同的田间管理措施。
图1
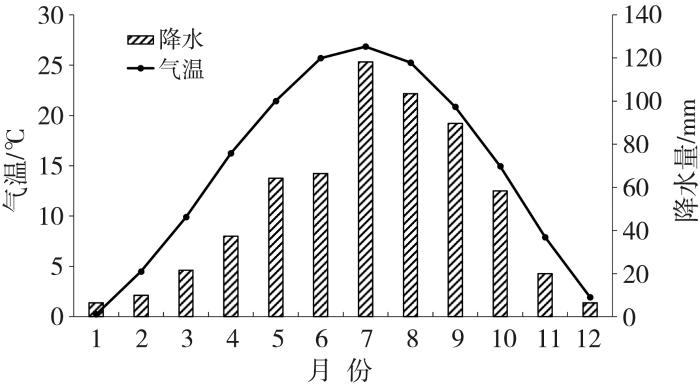
图1
1981—2020年试验区多年平均气温和降水量的逐月变化
Fig.1
The monthly variation of the average temperature and precipitation in the experimental area from 1981 to 2020
1.1.2 开顶式气室OTC
增温装置开顶式气室OTC为正六边形棱台,顶部敞口,气室无底,直接立于田间,其结构见图2。气室材料选用透光性≥98%的亚克力板。在OTC和CK内,均在30 cm处安装GM1366温度计记录温度。OTC设计高度为60 cm。
图2
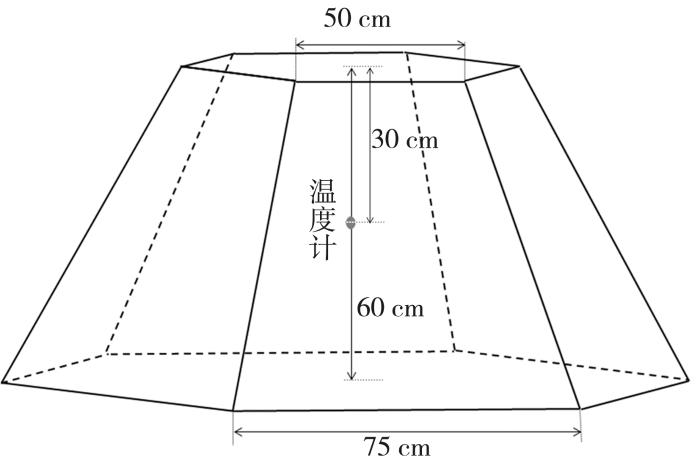
1.1.3 小麦种植与田间管理
在两个处理组播种前,施用黄腐三铵(N+P2O5+K2O≥36%)复合肥,施用量为749.6 kg·hm-2。供试小麦为郑麦1860号(Triticum aestivum L.,Zheng Mai 1860),该品种是当地主栽品种。小麦于2022年10月10日采用撒播方式播种,播种量224.9 kg·hm-2。返青期和灌浆期各灌溉一次,观测期间严格清除田间杂草。2023年6月6日收获,全生育期239 d。
1.2 小麦生态参数的测定
1.2.1 小麦株高与密度
冬小麦物候期记录依据气象行业标准《农业气象观测规范冬小麦》(全国农业气象标准化技术委员会,2015)。对照组选择6个测点,每个测点随机取10株(茎);在OTC中随机选取10株(茎)。在不同物候期对选定植株进行定株观测。小麦返青期,测量地面至植株叶片伸直后的最高叶尖的距离;拔节期,测量地面至最上部一片展开叶片的基部叶枕的距离;抽穗期,测量地面至穗顶(不包括芒长)的距离。每3天观测1次。在OTC中选择0.5 m×0.5 m区域,统计区域内的株数,并计算1 m2内的植株密度;对照组选择4个0.5 m×0.5 m测点,统计测点内的株数,以此计算对照组的密度。
1.2.2 叶面积和生物量
在各生育期,从OTC选取6株小麦样株,齐地面剪下,装入样品袋,使用精度为0.001 g的电子秤分别称取全样株、叶片、茎、穗的鲜重;将茎、穗装入纸信封,置于75 ℃烘箱中干燥至恒重后,分别称取干物质质量(生物量);叶片部分称鲜重后,平铺于A4纸上用扫描仪扫描图像,在ArcGIS中采用非监督分类法统计叶片面积A,并通过总叶片面积除以叶片数得到单片叶片面积,因扫描数据量较大,通过Python编程对图像进行批处理。随后将叶片装入纸质样品袋,在恒温干燥箱中以105 ℃杀青1 h,之后以75 ℃在烘箱中干燥12 h后称重,之后每1 h称重1次,直至相邻两次称量差小于5‰时停止烘干,记录干重W。对照组采用相同方法。植株含水率为鲜重与干重的比值。观测频率为每3天一次。
1.2.3 根系特征
小麦收割后,在每个OTC中选择5株(茎)小麦样株,对照组在6个测点内每点同样选取5株(茎)。将根系挖出(深度30 cm),装入0.5 mm孔径的尼龙网袋,用水冲洗至根系无泥土残留,捡出非根系物质,并剪去根系上部的茎。清洗后的根系放入根盘,加入纯净水,用德国Win-HIZO Pro 2008根系扫描仪采集图像,并通过Win-RHIZO 根系分析系统定量分析总根长、根系总表面积、根系总体积。之后将所有根系于75 ℃干燥至恒重,称取根系干重。
1.2.4 参数计算
比叶面积(Specific Leaf Area,SLA)计算公式(罗恬等,2022)如下:
干物质分配指数(Dry Matter Partition Index,DI)计算公式(刘凯强等,2021)如下:
式中:Bi、B分别为不同物候期的某器官干物质和总干物质质量,单位均为g。
1.2.5 数据处理
所有生态参数均取测点的平均值,使用SPSS 16.0进行数据统计分析,其中OTC内外温度差异采用配对t检验评价差异显著性水平;生态参数的平均数差异显著性检验采用One-Way ANOVA方法(p<0.05为显著,p<0.01为极显著)。使用Origin 8.0软件绘制图表。
2 结果与分析
2.1 OTC内外温度的差异
表1显示了OTC内外温度在不同生育阶段的差异。从整个生育期来看,除OTC4和OTC6外,其余气室内外的温度均表现出显著或极显著差异。返青—起身期,OTC2、OTC4的内外温度差异极显著,OTC1、OTC3内外温度差异显著,而OTC5和OTC6的内外温度差异不显著(p>0.05);拔节—扬花期,所有OTC的内外温度差异均不显著(p>0.05);灌浆—乳熟期,OTC2的内外温度差异极显著,OTC1、OTC3、OTC4、OTC5的内外温度差异显著,而OTC6的内外温度差异不显著。
表1 6个气室内温度与气象站记录的温度差异性检验
Tab.1
| 物候期 | 增温气室 | 内外温度平均值/℃ | 标准差/℃ | 显著性 (双尾) | 物候期 | 增温气室 | 内外温度平均值/℃ | 标准差/℃ | 显著性 (双尾) |
|---|---|---|---|---|---|---|---|---|---|
| 全生育期 | OTC1 | 14.86 | 9.31 | <0.001** | OTC1 | 16.00 | 4.87 | 0.667 | |
| OTC2 | 14.45 | 9.51 | 0.005** | OTC2 | 15.68 | 4.84 | 0.906 | ||
| OTC3 | 14.25 | 9.33 | 0.024* | OTC3 | 15.27 | 4.53 | 0.774 | ||
| OTC4 | 14.06 | 9.44 | 0.074 | 拔节—扬花期 | OTC4 | 15.20 | 4.59 | 0.721 | |
| OTC5 | 14.14 | 9.25 | 0.045 | OTC5 | 15.06 | 4.42 | 0.611 | ||
| OTC6 | 13.74 | 9.19 | 0.370 | OTC6 | 14.19 | 4.28 | 0.164 | ||
| 气象站 | 13.4 | 9.47 | 气象站 | 15.56 | 4.47 | ||||
| 返青—起身期 | OTC1 | 4.38 | 0.82 | 0.013* | OTC1 | 24.44 | 4.69 | 0.012* | |
| OTC2 | 3.12 | 0.61 | 0.007** | OTC2 | 24.54 | 4.68 | 0.007** | ||
| OTC3 | 3.20 | 0.60 | 0.045* | OTC3 | 24.24 | 4.71 | 0.029* | ||
| OTC4 | 2.99 | 0.56 | 0.000** | 灌浆—乳熟期 | OTC4 | 24.34 | 4.68 | 0.018* | |
| OTC5 | 3.31 | 0.57 | 0.353 | OTC5 | 24.20 | 4.70 | 0.038* | ||
| OTC6 | 3.21 | 0.55 | 0.111 | OTC6 | 23.93 | 4.85 | 0.125 | ||
| 气象站 | 3.38 | 4.27 | 气象站 | 23.07 | 4.81 |
注:*、**分别表示p<0.05差异显著、p<0.01差异极显著。
OTC6内外温度差异不显著可能与其位于试验地边缘,受到隔离带影响有关。为减少温度增温的不确定性,后续研究中可补充安装主动增温装置。
不同物候期气温变化幅度如图3所示。分蘖、返青和拔节期,OTC内温度增幅明显高于其他时期。整个生育期内,OTC1始终处于增温状态;OTC5、OTC6在抽穗、扬花和灌浆期气室内温度低于气室外;OTC2、OTC3和OTC4在抽穗、扬花和灌浆期气室内外温度差异较小;乳熟期,所有气室内温度均高于气室外,但内外增幅较小。总体上,冬小麦生育期内OTC1、OTC2、OTC3、OTC4、OTC5、OTC6气室温度的增幅分别为1.59、1.04、0.77、0.57、0.66和0.22 ℃,分别增大14.90%、9.75%、7.23%、5.36%、6.21%、2.07%。灌浆期典型5 d6个气室平均温度与气温温差变化(图4)表明,白天温度增加、夜间温度减小。07:30(北京时,下同),气室内外温差上升,13:00温差达到最大;16:00,气室内外温差变为负值,直至次日07:30温差再次转为正值。
图3
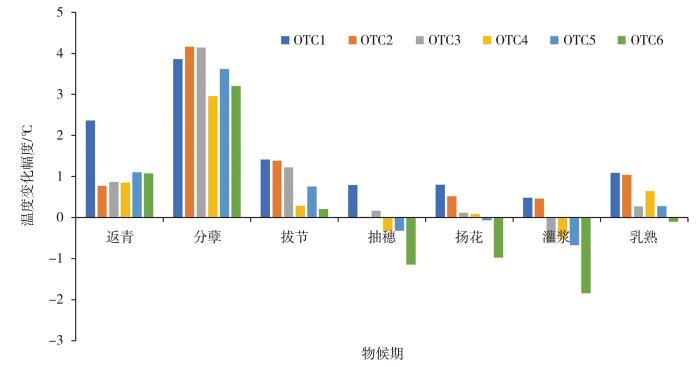
图3
不同物候期不同气室内外温度变化幅度
Fig.3
The variation amplitude of temperature inside and outside six OTCs during different phenological period
图4
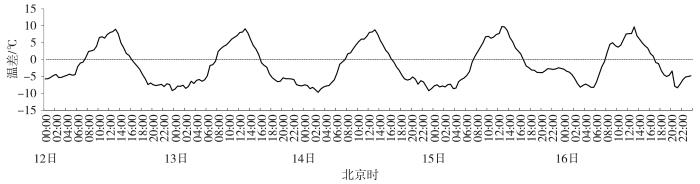
图4
灌浆期典型日(5月12—16日)气室平均温度与气温温差日变化
Fig.4
Diurnal variation of the temperature difference between average temperature of six OTCs and the air temperature on typical days (May 12 to 16) during grouting period
2.2 OTC内外生态参数对比
表2 观测期气室内外冬小麦生态参数对比
Tab.2
| 物候期 | 开始日期 | 株高/cm | 叶片面积/(cm2·片-1) | 比叶面积/(cm2·g-1) | 地上生物量/(g·m-2) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| OTC | CK | OTC | CK | OTC | CK | OTC | CK | OTC | CK | ||
| 返青 | 02-16 | 02-23 | 20.67Aa | 12.60Ab | 6.42a | 4.04b | |||||
| 起身 | 03-09 | 03-15 | 34.20Aa | 28.37Bb | 12.16Aa | 8.28Bb | 116.81Aa | 167.75Aa | 560.58Aa | 335.76Ab | |
| 拔节 | 03-29 | 04-08 | 61.67Aa | 52.04Bb | 16.87Aa | 15.17Ba | 247.70Aa | 242.27Aa | 678.35Aa | 511.16Bb | |
| 抽穗 | 04-21 | 04-26 | 76.94Aa | 66.15Bb | 19.44Aa | 20.18Aa | 219.07Aa | 220.44Ba | 1858.59Aa | 1031.78Bb | |
| 扬花 | 05-08 | 05-13 | 76.67Aa | 71.55Bb | 18.91Aa | 19.58Aa | 183.48Aa | 192.52Ba | 2847.39Aa | 1239.83Bb | |
| 灌浆 | 05-14 | 05-17 | 74.60Aa | 70.13Bb | 16.50Aa | 19.51Ab | 144.83Aa | 142.43Aa | 4434.99Aa | 2319.78Ab | |
| 乳熟 | 06-01 | 06-04 | 72.88Aa | 67.73Bb | 8.18Aa | 16.72Ab | 2885.84Aa | 1232.48Ab | |||
注:气室内各参数为6个气室平均;不同小写字母表示同一参数OTC和CK间差异显著(p<0.05),不同大写字母表示同一参数物候期间差异显著(p<0.05)。
图5
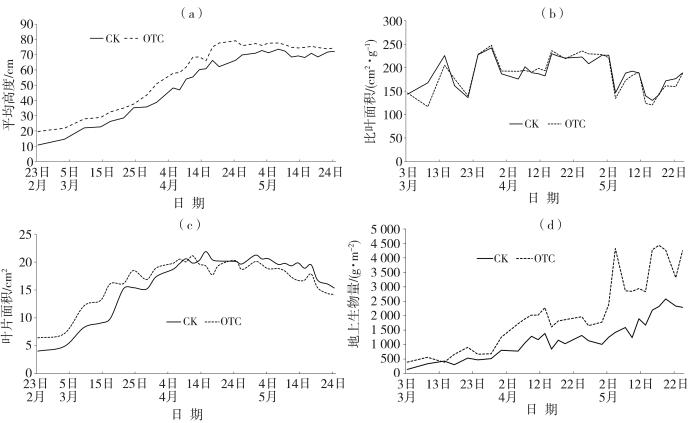
图5
2023年2月23日—5月24日(a、c)及3月3日—5月24日(b、d)OTC与对照组冬小麦平均高度(a)、比叶面积(b)、叶片面积(c)及地上生物量(d)变化
Fig.5
Daily variations of the average height (a), specific leaf area (b), leaf area (c), and aboveground biomass (d) of winter wheat in six OTCs and CK from February 23 to May 24 (a, c), and from March 3 to May 24 (b, d) 2023
冬小麦比叶面积在两种处理中随时间呈波动变化,但OTC与对照组差异不显著(p>0.05)。地上生物量随时间呈增加趋势,均在灌浆期达到峰值后下降。与对照组相比,OTC内地上生物量显著增加,尤其是拔节期后;而在抽穗、扬花、灌浆和乳熟期OTC内地上生物量分别增加80.13%、129.66%、91.18%、134.15%。整个生育期内,OTC与对照组冬小麦的地上生物量差异极显著。
综上所述,增温显著促进了冬小麦地上生物量的增长,特别是在返青期后。
2.3 根系形态特征的差异
细根(直径<2 mm)是根系的主要组成部分,在提高作物生产力和抗胁迫能力方面发挥重要作用(Luke Mccormack et al.,2015;Freschet et al.,2021)。本文仅统计直径<2 mm的根系形态特征。根据细根直径(L)大小,将根系分为4类:I类(0<L≤0.50 mm)、II类(0.50 mm<L≤1.00)、III类(1.00<L≤1.50)和IV类(1.50<L≤2.00)。从图6可看出,I类根系占比最大,其次是II类,两类根系长度占总根长的90%以上;I类、II类根系表面积占总根表面积的70%以上。不同OTC对根系的作用存在差异,总体而言,增温对细根长度具有促进作用,相比对照组,OTC组单株根系长度增加37.48%,单株细根表面积增加35.28%;但增温对根系总生物量有抑制作用,OTC组比对照组减少7.60%。OTC组与对照组单株根系根长和单株根总表面积均无显著差异(p>0.05)。
图6
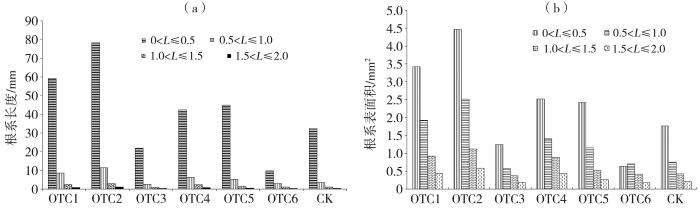
图6
6个OTC和对照组4类细根的根长(a)及根系表面积(b)
Fig.6
Root length (a) and root surface area (b) of four types of fine roots for six OTCs and CK
2.4 干物质累积及分配指数的差异
观测期内,两个处理组冬小麦叶、茎和穗的干物质累积量均呈递增趋势(表3),但不同器官在不同处理条件下达到最大值的物候期有所不同。对照组叶、茎、穗的干物质累积量分别在扬花、灌浆、乳熟期达到最大值;OTC组,叶的干物质累积量在孕穗期达到最大,后两者在灌浆期达到最大。对比两个处理组,对照组的叶、茎和穗的干重均低于OTC组,孕穗期叶的干物质累积量差异显著,而茎、穗在乳熟期无显著差异,其余物候期差异显著(p<0.05)。OTC组冬小麦叶干重在孕穗、扬花和灌浆期分别较对照组高58.24%、5.17%、7.48%;但在乳熟期OTC组叶干重较对照组小14.51%。两个处理组的茎干重均在灌浆期达到最大值,在乳熟期明显下降。与对照组相比,OTC对冬小麦茎干重有明显促进作用;对照组茎干重在孕穗、扬花、灌浆和乳熟期分别较OTC组减少97.26%、64.61%、92.08%、58.99%。穗干重方面,对照组在乳熟期达到峰值,OTC组在灌浆期达到最大值。在冬小麦整个生育期内,OTC组穗干重较对照组平均增加100.02%,说明增温处理对冬小麦穗干重也具有显著的促进作用。
表3 气室内外冬小麦干物质分配量对比
Tab.3
| 物候期 | 叶 | 茎 | 穗 | |||
|---|---|---|---|---|---|---|
| CK | OTC | CK | OTC | CK | OTC | |
| 孕穗 | 181.20a | 286.73b | 800.36a | 1578.81b | 326.98a | 786.20b |
| 扬花 | 204.22a | 214.67a | 946.44a | 1557.92b | 586.60a | 1124.05b |
| 灌浆 | 204.12a | 219.50a | 1048.82a | 2014.59b | 951.16a | 2011.36b |
| 乳熟 | 121.12a | 103.54a | 648.91a | 1031.73a | 1053.54a | 1649.17a |
注:不同字母代表不同物候期不同冬小麦组织在CK与OTC的显著差异(p<0.05)。
OTC内外冬小麦各器官在各物候期的干物质分配指数如图7所示。孕穗期,两个处理组茎干物质分配指数均较高,二者差异不大;但叶和穗的分配指数存在差异,对照组叶干物质分配比高于OTC组,而穗干物质分配指数反之。扬花期,茎干物质分配仍占主导地位,两个处理组茎干物质分配指数无明显差异;穗干物质分配比例均明显上升,而叶干物质分配指数较小,对照组叶干物质分配比例高于OTC组,而穗干物质分配指数对照组比OTC组低5.04%。灌浆期,两个处理组茎和穗干物质分配比例均较高,茎干物质分配指数两组无差别,穗干物质OTC组高于对照组;叶干物质分配指数较小,对照组叶干物质分配比例高于OTC组。乳熟期,穗干物质分配指数最大,在OTC组和对照组分别为57.77%、59.23%;茎干物质分配指数相近,分别为35.58%、37.05%;对照组叶干物质分配比例高于OTC组,但两组中分配指数均较低。从孕穗期到乳熟期,随着生育进程推进,叶和茎的干物质分配指数逐渐下降,穗的干物质分配指数明显上升。
图7
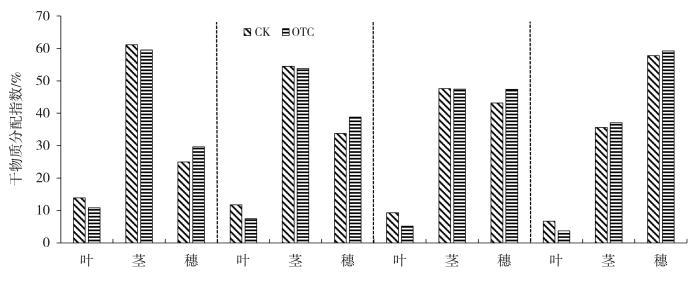
图7
不同物候期冬小麦不同器官干物质分配指数
Fig.7
Comparison of dry matter allocation index of winter wheat inside and outside OTC at different phenological periods
3 讨论
3.1 增温设施的气室效应
大量研究表明,OTC具有明显的气室效应,能够对温度、湿度、风速和光照等环境因子产生影响,尤其是使内部温度升高,其增温幅度为0~8.0 ℃(Hogsett et al.,1985;Sanders et al.,1990;Leadley and Drake,1993;Fuhrer,1994;曹嘉晨等,2018)。本研究结果显示,实验期间OTC整体平均增温0.8 ℃。田间热成像监测结果进一步表明,6个OTC区域地表均有明显的增温效果(图8)。OTC的增温效应受环境温度、时间及物候期等因素的影响。气温较低时,OTC的温度增幅较大(Mcleod and Long,1999)。作物生长初期的增温幅度较高,而在后期,因作物的群体遮蔽,导致冠幅下部对增温响应慢且增幅较小(陈金等,2010);08:00—18:00,OTC内温度高于外部环境温度,而18:00—次日08:00则相反(童思思等,2023),这些都与本文研究结果相似,仅增温与降温的时间节点有所差异。OTC内温度的增幅具有明显季节性差异,冬春季明显高于夏秋季(房世波等,2010)。总之,OTC增温效应显著,符合气候变暖相关农田试验研究的要求,并具有简单、经济、可靠及可保持作物原位生长等优点(童思思等,2023)。值得注意的是,OTC除了增加内环境温度之外,也会对湿度、光强和风速造成影响(邱秋金等,2008),从而影响作物生长。总之,增温设施(OTC)具有显著的增温效应,但增温效果受环境因素及作物发育阶段的影响。
图8
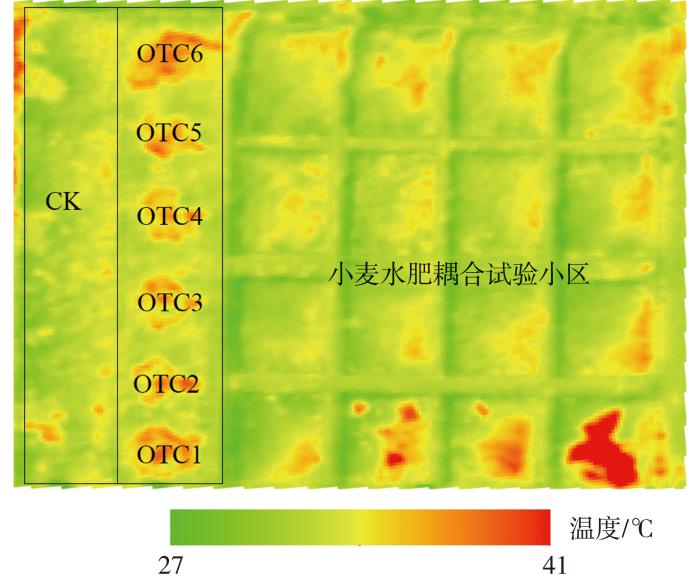
图8
2023年6月4日10:34:26 OTC与对照组冬小麦地表热成像图
Fig.8
Surface thermal image in winter wheat field for OTC and CK
3.2 增温对冬小麦地上功能性状的影响
研究发现,OTC通常能够促进植物株高的增加(Sanders et al.,1990;李白玉等,2021),本研究也得到类似结论。然而,曹嘉晨等(2018)也指出,OTC在某些情况下可能导致株高降低。一般而言,日平均温度越高,叶面积指数越大(郭玉龙等,2023),因此OTC常导致植物叶面积增大(曹嘉晨等,2018;刘国一等,2018),但本研究显示,抽穗期后OTC对冬小麦叶片面积具有抑制作用,这可能与OTC内发生的叶锈病有关。叶锈病在抽穗期后造成叶片停止生长、枯死或内卷,其发生与OTC的增温效应密切相关。暖冬时锈病菌潜育繁殖明显(张淑玲等,2005),叶锈病在孕穗至抽穗期迅速发展(彭超和刘丽然,2001),春小麦锈病发病率随温度升高明显增加(王鹤龄等,2015)。
本研究表明,增温对冬小麦的影响具有阶段性:营养生长期,增温明显促进冬小麦生长;生殖生长期,增温对冬小麦的促进作用不显著,甚至表现为抑制作用。OTC的增温效应在一定程度上加剧了叶锈病的发生,这不仅影响了冬小麦叶片的光合作用和物质积累,还对整个生育期的生长发育及产量形成产生了负面作用。因此,未来研究应进一步探讨增温条件下叶锈病发生的具体机制及其控制策略,以更全面地评估OTC实验的适用性。
3.3 增温对冬小麦地下功能性状的影响
3.4 增温对冬小麦干物质量分配的影响
研究表明增温设施(OTC)增温效应明显。增温对小麦的影响呈阶段性变化,营养生长期,增温明显促进小麦生长;生殖生长期,增温对小麦的生长发育促进作用不显著,甚至表现为抑制作用。此外,增温还明显促进了叶锈病的发生。总体而言,本研究表明增温不仅对冬小麦的干物质积累和分配产生重要影响,还可能通过调节生物量分配,帮助作物应对温度变化引起的水分胁迫。未来的研究可以进一步探讨增温对作物生长的长期影响以及其他环境因素的相互作用,为应对气候变化提供更有效的农业管理建议。
参考文献
晚冬早春阶段性增温调控华北平原北部小麦源库性能的作用
[J].
针对华北平原北部冬小麦生长发育所需温度与实际环境温度间的矛盾,于2019-2021年连续2个生长季,通过大田试验研究了晚冬早春阶段性升温调控小麦源库性能的作用。首个生长季设置4个阶段性升温处理:1月20日(CT1)、1月26日(CT2)、2月1日(CT3)、2月7日(CT4)增温,3月20日结束增温;第2个生长季设置3个阶段性升温处理:1月25日(CT1)、2月1日(CT2)、2月8日(CT3)增温,3月15日结束增温;2个生长季均以常规生产为对照(CK)。结果表明:增温处理增温阶段积温增加138.1~405.1 ℃,第二生长季CT1拔节-开花日均温降低2.50 ℃,第一生长季开花-成熟日均温降低2.31 ℃,第一生长季提前小麦返青25 d,且延长返青-成熟总天数21 d。第二生长季CT1开花期叶面积指数可显著提高17.6%,旗叶面积可显著提高33.7%。2020-2021生长季花后5 d净光合速率可显著提高11.7%,第一生长季花后CT1旗叶丙二醛含量可显著下降28.0%。在第二生长季中,CT1穗长可显著提高15.7%,粒长显著提高2.3%,花后15 d籽粒灌浆速率则可显著提高41.0%,CT1穗粒数可显著提高8.8粒,千粒质量显著提高2.0 g,产量可显著提高35.8%。由此表明,增温处理提前了小麦返青,小麦源库物质积累的开始时间提前,结束增温措施之后的相对降温,既延长了源库物质积累的总时间,同时又为源库活性的提高准备了条件,且存在增温处理实施时间越早,小麦源库性能提高越多的趋势。
干旱胁迫对‘青燕1号’燕麦产量及干物质积累与分配的影响
[J].为探究‘青燕1号’燕麦对不同干旱胁迫方式的响应变化,在盆栽试验条件下,采用不同干旱胁迫程度和胁迫次数处理,研究燕麦各器官干物质积累与分配及产量和产量因子的变化规律。结果显示:燕麦穗长、小穗粒数、单序籽粒重、百粒重和产量下降明显,小穗数、穗粒数有增有减,而空铃数无明显变化;干旱胁迫抑制了器官干物质积累,其中分配比例以茎和根部转移相对较多,穗部较少。从整体水平来看,不同胁迫程度和胁迫次数影响大小表现为:3次、中度和重度胁迫影响较大;不同胁迫时期变化下以苗期-拔节期、孕穗-抽穗期、开花-乳熟期干旱(SM)时期和苗期-拔节期、孕穗-抽穗期干旱(SH)影响最大。不同干旱胁迫下各指标相关性分析及通径分析得出,燕麦小穗数、穗粒数、小穗粒数、单序籽粒重、百粒重、穗长、穗干重、茎干重、根干重、叶干重、穗分配指数与产量呈显著正相关关系,相关系数在0.368~0.922,而茎分配指数、根分配指数、叶分配指数与产量呈负相关关系,相关系数在-0.673~-0.299,空铃数与产量为负相关关系,但未达到显著水平,为-0.021;通径分析发现,单序籽粒重、百粒重、穗粒数对燕麦产量增产具有重要作用。
植物功能性状与环境和生态系统功能
[J].植物性状反映了植物对生长环境的响应和适应,将环境、植物个体和生态系统结构、 过程与功能联系起来(所谓的“植物功能性状”)。该文介绍了植物功能性状的分类体系,综述了国内外植物功能性状与气候(包括气温、降水、光照)、地理空间变异(包括地形地 貌、生态梯度、海拔)、营养、干扰(包括火灾、放牧、生物入侵、土地利用)等环境因素,以及与生态系统功能之间关系的研究进展,探讨了全球变化(气候变化和CO<sub>2</sub>浓度升高 ) 对个体和群落植物功能性状的影响。植物功能性状的研究已经取得很多成果,并应用于全球变化、古植被恢复和古气候定量重建、环境监测与评价、生态保护和恢复等研究中,但大尺度、多生境因子下的植物功能性状研究仍有待于加强,同时需要改进性状的测量手段;我国 的植物功能性状研究还需要更加明朗化和系统化。
气候变化科学与人类可持续发展
[J].政府间气候变化专门委员会(IPCC)自2007年发布第四次评估报告(AR4)以来,新的观测证据进一步证明,全球气候系统变暖是毋庸置疑的事实。2012年之前的3个连续10年的全球地表平均气温,都比1850年以来任何一个10年更高,且可能是过去1400年来最热的30年。虽然1998-2012年全球地表增温速率趋缓,但还不能反映出气候变化的长期趋势。1970年以来海洋在变暖,海洋上层75 m以上的海水温度每10年升温幅度超过0.11℃;1971-2010年地球气候系统增加的净能量中,93%被海洋吸收。全球平均海平面上升速率加快,1993-2010年间高达3.2 mm/年。全球海洋的人为碳库很可能已增加,导致海洋表层水酸化。1971年以来,全球几乎所有冰川、格陵兰冰盖和南极冰盖的冰量都在损失。其中1979年以来北极海冰范围以每10年3.5%~4.1%的速率缩小,同期南极海冰范围以每10年1.2%~1.8%的速率增大。北半球积雪范围在缩小。20世纪80年代初以来,大多数地区的多年冻土温度升高。已在大气和海洋变暖、水循环变化、冰冻圈退缩、海平面上升和极端气候事件的变化中检测到人类活动影响的信号。1750年以来大气CO<sub>2</sub>浓度的增加是人为辐射强迫增加的主因,导致20世纪50年代以来50%以上的全球气候变暖,其信度超过95%。采用CMIP5模式和典型浓度路径(RCPs),预估本世纪末全球地表平均气温将继续升高,热浪、强降水等极端事件的发生频率将增加,降水将呈现“干者愈干、湿者愈湿”趋势。海洋上层的温度比1986-2005年间升高0.6~2.0℃,热量将从海表传向深海,并影响大洋环流,2100年海平面将上升0.26~0.82 m。冰冻圈将继续变暖。为控制气候变暖,人类需要减少温室气体排放。如果较工业化之前的温升达到2℃,全球年均经济损失将达到收入的0.2%~2.0%,并造成大范围不可逆的影响,导致死亡、疾病、食品安全、内陆洪涝、农村饮水和灌溉困难等问题,影响人类安全。但如果采取积极行动,2℃的温升目标仍可望达到。为遏制逐渐失控的全球变暖,需全球共同努力减排,以实现人类可持续发展的理想。
增温和降水变化对西北半干旱区春小麦产量和品质的影响
[J].为了明确未来气候变化对半干旱区春小麦产量和品质的影响,本研究选择典型半干旱区定西试验基地,利用开放式红外增温模拟系统和水分控制装置设置不同降水量(减少20%、不变、增加20%)和温度梯度\[大气温度(对照)、增加1.0 ℃、增加2.0 ℃、增加3.0 ℃\],模拟气候变化对春小麦生长发育、产量及其构成因素和品质的影响.结果表明:当增温幅度小于2 ℃时,降水变化对春小麦穗粒数影响不显著;当增温为3 ℃时,降水减少显著减少穗粒数,降水增加显著增加穗粒数.随着气温升高,降水减少对春小麦千粒重的负效应增大,春小麦不孕小穗数与气温呈二次曲线上升.降水减少20%条件下,增温1、2和3 ℃的春小麦产量分别下降12.1%、24.7%、42.7%;降水不变条件下,春小麦产量分别下降8.4%、15.1%、21.8%;降水增加20%条件下,春小麦产量分别下降9.0%、15.5%、22.2%.春小麦籽粒淀粉含量随温度的增加而下降,籽粒蛋白质含量随温度的增加而上升.增温2 ℃有利于蚜虫暴发,但增温3 ℃抑制蚜虫暴发;春小麦锈病发病率随着温度增加而上升.
世界粮食安全面临新的严峻挑战
[J].
Effect of heat stress on the growth, root sugars, acid invertase and protein profile of pepper seedlings following transplanting
[J].
Root growth dependence on soil temperature for Opuntia ficus-indica: Influences of air temperature and a doubled CO2 concentration
[J].
Root traits as drivers of plant and ecosystem functioning: Current understanding, pitfalls and future research needs
[J].
Effects of ozone on managed pasture: I. effects of open-top chambers on microclimate, ozone flux, and plant growth
[J].Open-top chambers (OTC) were established in a field of managed pasture, and environmental parameters were recorded inside and outside to study the influence of OTCs on radiation, air temperature (T(air)), saturation vapour pressure deficit (svpd), and soil water content in relationship to plant growth and yield. Canopy development in OTCs supplied with non-filtered air (NF) and in ambient (AA) plots was followed by measuring leaf area index (LAI). The dry matter yield was determined after three growth periods in each of two consecutive seasons. Boundary layer conductance (g(bw)) and wind speed (u) were measured along a vertical profile, and day-time flux were measured along a vertical profile, and day-time flux of O(3) was estimated throughout the experiment on the basis of a mass balance. The vertical profile of u showed values in the range 1-1.2 m s(-1) at the top of the canopy, and maximum g(bw) was 20-25 mm s(-1). Average reduction in global radiation in OTCs was 25%, and volumetric soil water content was reduced by about 5%. Daily mean T(air) was increased by 1.3 degrees C, mean daily maximum svpd by 0.08 kPa, and the temperature sum (degree days with base temperature of +5 degrees C) by 12%. Fluctuations in the difference in daily mean T(air) and svpd during the daytime between OTCs and ambient air were related to canopy structure. Differences were largest after each cut and declined with increasing LAI. A small effect of changes in LAI on T(air) and svpd occurred during periods with low soil water content. The flux of O(3) in OTCs was largest (>100 microg m(-2) min(-1)) before and smallest (<20 microg m(-2) min(-1)) after each cut. Calculated deposition velocities for O(3) (nu(d)) in the range 0-3 cm s(-1) were generally higher than those measured under most field conditions. Overall, in OTCs the deficit in soil and atmospheric moisture was larger than in the open field, and the increase in daily mean T(air) was strongly influenced by the stage of canopy development. Changes in microclimate and incoming radiation affected pasture development. LAI was slightly reduced in OTCs as compared to AA plots. The total accumulated dry matter yield for all six growth periods was only about 7% lower in OTCs, but the contribution of clover to total forage mass declined during the experiment. OTCs had no significant effect on weeds. The results indicate that OTCs reduced the competitiveness of clover, and that the increase in growth of grasses compensates for the loss in clover yield. The shift in species composition caused by OTCs must be considered when studying the effect of pollutants on pasture.
Impacts of recent climate warming, cultivar changes, and crop management on winter wheat phenology across the Loess Plateau of China
[J].
A programmable exposure control system for determination of the effects of pollutant exposure regimes on plant growth
[J].
Effects of field experimental warming on wheat root distribution under conventional tillage and no-tillage systems
[J].
Root carbon and protein metabolism associated with heat tolerance
[J].
Open top chambers for exposing plant canopies to elevated CO2 concentration and for measuring net gas exchange
[J].
Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes
[J].Fine roots acquire essential soil resources and mediate biogeochemical cycling in terrestrial ecosystems. Estimates of carbon and nutrient allocation to build and maintain these structures remain uncertain because of the challenges of consistently measuring and interpreting fine-root systems. Traditionally, fine roots have been defined as all roots ≤ 2 mm in diameter, yet it is now recognized that this approach fails to capture the diversity of form and function observed among fine-root orders. Here, we demonstrate how order-based and functional classification frameworks improve our understanding of dynamic root processes in ecosystems dominated by perennial plants. In these frameworks, fine roots are either separated into individual root orders or functionally defined into a shorter-lived absorptive pool and a longer-lived transport fine-root pool. Using these frameworks, we estimate that fine-root production and turnover represent 22% of terrestrial net primary production globally - a c. 30% reduction from previous estimates assuming a single fine-root pool. Future work developing tools to rapidly differentiate functional fine-root classes, explicit incorporation of mycorrhizal fungi into fine-root studies, and wider adoption of a two-pool approach to model fine roots provide opportunities to better understand below-ground processes in the terrestrial biosphere.© 2015 The Authors. New Phytologist © 2015 New Phytologist Trust.
Free-air carbon dioxide enrichment (FACE) in global change research: A review
[J].
Dynamic adaptation of maize and wheat production to climate change
[J].
The growth and development of Vicia faba L. in filtered and unfiltered open-top chambers
[J].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}