

0 引言
作物生长模拟模型(简称“作物模型”)通过考虑影响作物生长的各种因素来描述作物生育周期内的形态与生理指标,通常表现为一种数学公式(李翠云等,2022;蒙继华等,2024)。早期关于设施作物模型的研究主要以温度为基础来定义发育周期(Heuvelink,1996),如基于日平均温度函数模拟番茄果实生长发育(Heuvelink,1999)、基于“源库”理论模拟设施作物的干物质分配(Marcelis,1993a;Marcelis,1994)等。这些建模方法虽然机理性强,但在模型精确度与灵活度上存在不足。后续研究表明,相较于机理性模型,经验性模型在作物模拟方面的精确度更高(Marcelis,1993b)。如Heuvelink(1995)通过对番茄进行了12 a的破坏性试验,发现番茄的成熟日期、叶面积生长、果实分配比等生长指标受光照和温度的共同控制,并与之呈正相关关系。Baker et al.(2001)进一步研究发现,仅使用温度来预测设施作物的生长会导致模拟产量低于实际产量,因为除了温度外,设施果实的生长还与光合有效辐射(Photosynthetically Active Radiation,PAR)有关,适当提高生育期间的光合有效辐射和温度可有效提高番茄果实的干物质质量(Jones,2017)。这些研究表明,温度和光照是设施作物生长不可或缺的环境因素。为量化温度和光照对设施作物的影响,引入Logistic模型来描述作物的非线性生长轨迹,该模型能有效反映环境变化对作物生长的动态影响(Caloin and Yu,1982),被广泛应用于模拟设施作物的生长与干物质分配。利用该模型模拟设施番茄的生长发育和果茎增长,结果显示模型具有良好的适用性和预测性(He and Zhang,2006;Fang et al.,2022)。为进一步提高模型精确度,研究人员引入了辐热积概念(倪纪恒等,2006)。辐热积法可量化光合有效辐射和温度热效应,通过将设施作物的生长指标与辐热积建立Logistic模型关系,可准确模拟诸如芹菜(程陈等,2021)、番茄(王丹丹等,2018)、烤烟(张明达等,2013)等作物的外观形态与干物质分配,并系统化地表现出设施蔬果的生长动态(程陈等,2021)。相较积温法,使用辐热积法建立的Logistic模型精确度更高,如可将黄瓜叶面积指数的精度提高37%(李永秀等,2006;倪纪恒等,2009)。因此,基于辐热积的Logistic模型精确度高、实用性强、普适性好,对设施作物的模拟研究具有重要价值。
以往研究设施番茄的Logistic模型主要聚焦于干物质分配,而对番茄的其他生理指标探讨较少。然而,设施番茄最终产量实际受生育期内多个生理指标的共同影响。因此,本文基于辐热积模型拟合番茄开花坐果数、果茎生长量与干物质等关键生长指标,构建综合的番茄生长Logistic曲线模型并进行验证,以期为帮助种植户合理调控温室环境、提高番茄产量、规避灾害风险,并为预测番茄的产量品质与上市时间提供理论支持。
1 材料与方法
1.1 试验设计
试验分别在山东省临沂市、济南市和济宁市开展,各实验点的大棚均呈东西走向、坐北朝南,覆盖阿尔法薄膜有限公司生产的聚乙烯无滴膜,透光系数为75%,供试土壤类型均为黏壤土。试验环境见表1。
表1 试验环境
Tab.1
| 试验 | 地点 | 经纬度 | 试验品种 | 定植日期 | 试验日期 | 结束日期 | 垄宽/cm | 行间距/cm | 株间距/cm |
|---|---|---|---|---|---|---|---|---|---|
| 试验1 | 临沂市沂南县设施农业气象试验站 | 118.47°E,35.56°N | 粉冠 | 2017年10月1日 | 2017年11月10日 | 2018年1月1日 | 95 | 40 | 30 |
| 试验2 | 济南市长清区济西农业设施蔬菜大棚 | 116.76°E,35.56°N | 粉冠 | 2018年10年10日 | 2018年11月20日 | 2019年1月10日 | 60 | 40 | 30 |
| 试验3 | 济宁市兖州瑞鹏农业设施蔬菜大棚 | 116.75°E,35.56°N | 普罗旺斯 | 2021年10月7日 | 2021年11月17日 | 2022年1月3日 | 50 | 40 | 20 |
日光温室大棚聚光保温效果好,棚内温度能始终保持在10~30 ℃,适宜设施作物生长。为避免自然天气状况变化对试验结果的影响,使用环境检测仪监控棚内光照、温度与湿度的实时变化。遇到长时间低温寡照时,及时人工补光,维持足够光照;在阴雨天气及夜间,通过增加炭盆和棚顶覆盖棉被等措施保温;晴好天气时,进行午间通风降湿。根据番茄不同生育期和土壤墒情状况进行灌溉,以保持土壤湿度在适宜设施番茄生长的水平(朱雨晴和薛晓萍,2020a;朱雨晴和薛晓萍,2020b)。按照高产栽培标准进行田间管理,苗期施1次高氮肥,用量1.5 g·m-2,幼果期(90%的植株开始结果后第10 d)开始施用氮磷钾(1:1:1)肥,用量7.5 g·m-2,此后每15 d施一次同样比例和用量的肥料。根据天气状况,温室大棚的揭棚时间为9:00—10:00,盖棚时间为16:00—17:00,使试验期内10:00—16:00的平均光合有效辐射为400~700 μmol·m-2·s-1,日平均气温为15~25 ℃,日平均空气相对湿度为75%~85%(图1)。
图1
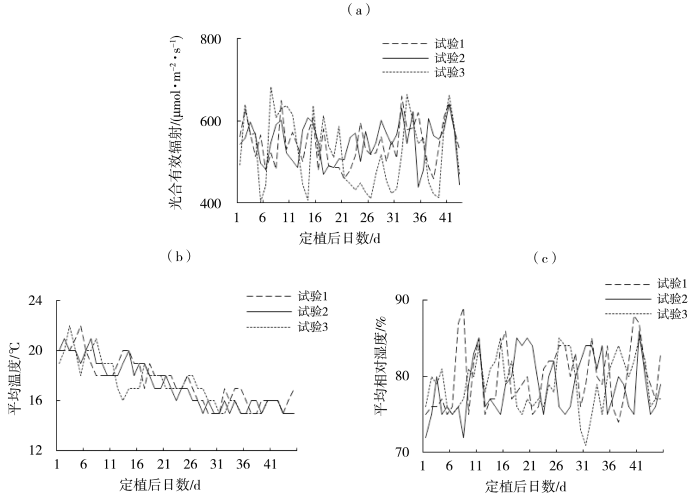
图1
三个试验点番茄生育期内平均光合有效辐射(a)、日平均温度(b)和日平均相对湿度(c)的变化
Fig.1
Changes in average PAR (a), daily average temperature (b) and daily average relative humidity (c) during the experiment
1.2 气象数据来源与处理
采用英国制造的WS-GP2小型自动气象站测定3个日光温室内的光合有效辐射,气温、空气相对湿度数据通过美国制造的WatchDog 2000数据采集器获得。数据采集频率为每5 min一次,计算每小时的平均值以得到逐时光合有效辐射、平均气温和平均相对湿度。使用Excel 2010软件进行数据整理,建立相关数据表格、绘制折线图和散点图以进行数据分析和可视化。应用SPSS 26.0软件确定Logistic曲线模型的参数。
1.3 测定项目
1.3.1 番茄开花坐果数
在3个站点的番茄温室试验中,整个番茄生长期间,每个站点选择50株健康且生长状态相似的番茄植株作为观测样本,对每株番茄挂牌进行定株观测。试验前,所有选定植株的花序开花数记为0,最大花蕾长度设为0.5 cm。试验开始后,每3 d记录一次植株的开花数和坐果数,直到这些数值不再发生变化时停止记录。开花的标准为花瓣展开角度达到45°,坐果的标准为果实横茎达到10 mm(朱雨晴和薛晓萍,2020a)。
1.3.2 番茄果茎生长
选择50株与开花坐果期不同但生长状态相似的健康番茄植株作为样本,对每株番茄挂牌进行定株观测。标记坐果日期相同、形态相似的健康果实,定期跟踪观测。自坐果之日起,每3 d使用精度为0.01 mm,量程为150 mm的公英制转换500系列CD-AX数显卡尺测量一次果实的横茎和纵茎尺寸,其中横茎为果实最宽处长度,纵茎为从番茄柄到果实顶点的长度(朱雨晴和薛晓萍,2020b)。
1.3.3 番茄干物质
在试验中,每3 d选择10株未挂牌且生长状态相似的健康番茄植株进行破坏性取样,整个试验期间共取样20次,总样本数为200株。取样后,将每个植株细分为茎叶、花果和根茎3部分。所有部分清水洗净后擦干,分别装入牛皮纸袋中,放入烘干机在105 ℃下杀青30 min,随后在85 ℃下烘干至恒定质量。之后,每次测量10株植株的干物质质量并记录平均值。番茄茎叶干物质量、花果干物质量和根干物质量的分配比分别为番茄茎叶干物质量、花果干物质量和根干物质量占植株总干物质量的比值。
1.4 生长模型构建
1.4.1 辐热积模型
式中:To、Tf、Tm、T(℃)分别为番茄生育期内最适生长温度、生长下限温度、生长上限温度、试验环境中每小时平均温度。番茄不同生育期的三基点温度参考倪纪恒等(2006)的研究。
表2 番茄各生育期三基点温度
Tab.2
| 不同生育期 | 下限温度Tf | 最适温度To | 上限温度Tm |
|---|---|---|---|
| 苗期 | 10 | 25 | 30 |
| 花期 | 15 | 25 | 30 |
| 结果期 | 15 | 25 | 35 |
| 采收期 | 15 | 25 | 35 |
1.4.2 番茄生长模型
番茄开花坐果数、果茎生长量与干物质积累的动态生长变化符合Logistic曲线模型,Logistic曲线模型的公式如下:
式中:y为番茄试验观测的测定值,如开花数(朵)、坐果数(个)、果茎长宽(mm)、干物质分配比和干物质积累量(g);x(mol·m-2)为累计辐热积;k、a、b均为模型参数,由SPSS软件拟合得到;e为自然常数2.718。
对Logistic曲线模型的公式分别求一阶、二阶和三阶导数(薛晓萍,2007),得到番茄在辐热积累积到一定程度后的始盛点(p1)、盛末点(p2)和高峰点(pm),单位均为mol·m-2,公式如下:
生长速率计算公式为:
最大相对生长速率公式为:
1.4.3 模型验证方法
1.4.4 模型验证
为验证设施番茄生长发育的Logistic曲线模型的精确性和普适性,以试验1第二茬花序的植株为基础,建立Logistic曲线模型后,选择试验1第三茬花序作为验证1,验证不同茬口对Logistic曲线模型的适用性(干物质验证时不分花序,下同);选择试验2(3)第二茬花序作为验证2(3),验证不同种植地区相同(不同)品种对Logistic曲线模型的适用性。
2 结果与分析
2.1 模型模拟结果
图2
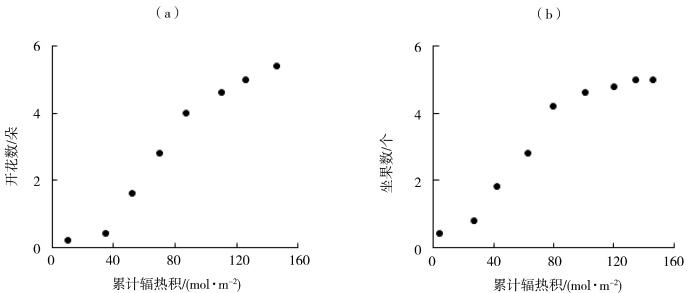
图2
番茄开花数(a)、坐果数(b)随累计辐热积的变化
Fig.2
The changes of the number of tomato flowering (a) and fruit setting (b) with cumulative PTP
图3
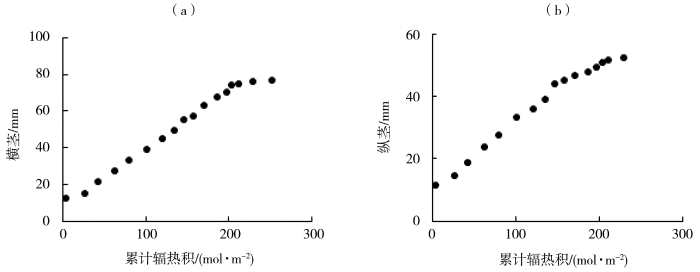
图3
番茄果实横径(a)、纵茎(b)随累计辐热积的变化
Fig.3
The changes of tomato fruit width (a) and length (b) with cumulative PTP
图4
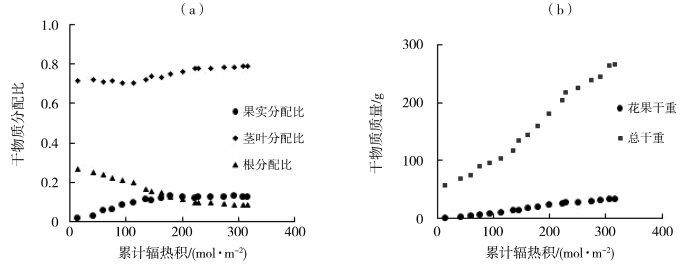
图4
番茄干物质分配比(a)、干物质量(b)随累计辐热积的变化
Fig.4
The changes of the dry matter partitioning and dry matter accumulation of tomato with cumulative PTP
表3 番茄生育指标的生长模型与特征值
Tab.3
| 生育指标 | 生长模型与特征值 | ||||||
|---|---|---|---|---|---|---|---|
| 生长模型 | R2 | p1/(mol·m-2) | p2/(mol·m-2) | p3/(mol·m-2) | Vmax | ||
| 编号 | 公式 | ||||||
| 开花 | (6) | y=5.238/[1+3.979exp (-0.058x)] | 0.994 | 1.10 | 46.52 | 23.81 | 0.08朵·d-1 |
| 坐果 | (7) | y=5.028/[1+2.995exp (-0.054x)] | 0.994 | 0 | 44.70 | 20.31 | 0.07个·d-1 |
| 果实横茎 | (8) | y=87.782/[1+1.888exp (-0.016x)] | 0.996 | 0 | 122.03 | 39.72 | 0.35 mm·d-1 |
| 果实纵茎 | (9) | y=60.573/[1+1.477exp (-0.016x)] | 0.995 | 0 | 106.69 | 24.38 | 0.24 mm·d-1 |
| 果实分配比 | (10) | y=0.126/[1+2.436exp (-0.034x)] | 0.992 | 0 | 64.92 | 26.19 | 0 |
| 花果干物质量 | (11) | y=30.74/(1+3.066exp (-0.021x)) | 0.998 | 0 | 116.07 | 53.35 | 0.16 g·d-1 |
| 总干物质量 | (12) | y=366.158/(1+1.982exp (-0.011x)) | 0.998 | 0 | 181.92 | 62.19 | 1.01 g·d-1 |
图2为试验1中番茄开花数、坐果数的观测值与累计辐热积的关系。可以看出,番茄开花数、坐果数与累计辐热积符合“S”型生长曲线。生长速率经历了“慢、快、慢”的变化,最终趋于平稳。番茄开花数达到最大时所需的累计辐热积为146.59 mol·m-2,坐果数达到最大时所需的累计辐热积为146.90 mol·m-2。番茄开花数、坐果数与累计辐热积之间的Logistic模型决定系数(R2)均为0.994。累计辐热积为1.10~46.52 mol·m-2时,番茄开花数迅速增长,其中辐热积为23.81 mol·m-2时开花速率最大,达0.08 朵·d-1;番茄坐果数在累计辐热积为0~44.70 mol·m-2时迅速增长,其中辐热积为20.31 mol·m-2时坐果速率达到最大,为0.07 个·d-1。
图3为试验1番茄果实横茎、纵茎的观测值与累计辐热积的关系,由图可知,番茄果实的横、纵径与累计辐热积符合“S”型生长曲线。番茄果实横茎达到最大时所需的累计辐热积为252.00 mol·m-2,纵茎达到最大时所需的累计辐热积为230.00 mol·m-2。由表3可知,番茄果实横径、纵径与累计辐热积之间的Logistic模型决定系数分别为0.996、0.995。累计辐热积在0~122.03 mol·m-2时为番茄果实横茎的快速生长阶段,其中在辐热积为39.72 mol·m-2时果实横茎的生长速率最大,为0.35 mm·d-1;累计辐热积在0~106.69 mol·m-2时为番茄果实纵茎的迅速生长阶段,在辐热积为24.38 mol·m-2时果实纵茎的生长速率最大,为0.24 mm·d-1。
图4为试验1的番茄干物质分配比、干物质积累量与辐热积的关系。番茄茎叶分配比、根分配比与辐热积的关系公式中存在无效系数,表明番茄的茎叶与根的分配比均不符合Logistic生长模型。果实分配比、花果干物质累积、总干物质累积与累计辐热积之间的Logistic拟合度均达到极显著水平;累计辐热积在0~64.92 mol·m-2时,番茄果实分配比快速增加,在辐热积为26.19 mol·m-2时分配比增加速率最大;辐热积在0~181.92 mol·m-2时,番茄总干物质量进入快速增长时期,累计辐热积为62.19 mol·m-2时,番茄总干物质量增长速率最大,为1.01 g·d-1;累计辐热积在0~116.07 mol·m-2时番茄花果干物质量迅速增长,辐热积为53.35 mol·m-2时,番茄花果干物质量增长速率最大,为0.16 g·d-1。
2.2 模型检验
利用与建立的模型独立的生长指标,输入环境数据得到番茄开花坐果数、果实横纵茎生长量、果实分配比、总干物质量及果实干物质量的模拟值,并与实测值进行对比(图5~7)。同时计算模拟值与实测值的R2、RMSE和nRMSE(表4),番茄开花坐果数、果茎生长量、果实分配比与干物质重的模拟值与实测值的R2为0.905~0.999,RMSE为0.208~14.229,nRMSE为0.027~0.246,整体精度较高,拟合效果较好。具体而言,第1组数据验证中,番茄开花数、坐果数的模型精确度较高,果实横、纵茎生长量的模型精确度极高;第2组数据验证中,番茄开花数模型精确度中等,坐果数和果茎生长量、以及果实干物质量和总干物质量的模型精确度极高,果实分配比的模型精确度较高;第3组数据验证中,番茄开花数、果实分配比和果实干物质量的模型精确度中等,坐果数和果茎生长量模型精确度较高,总干物质量的模型精确度极高。
图5
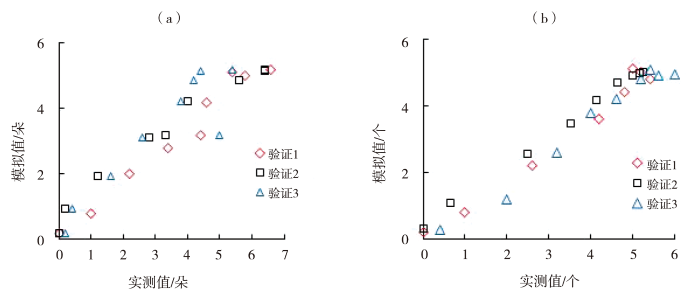
图5
番茄开花数(a)、坐果数(b)的模拟与实测散点图
Fig.5
Scatter graph of simulated and measured numbers of tomato flowering (a) and fruit sitting (b)
图6
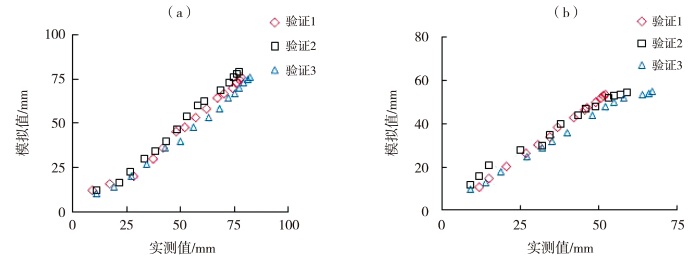
图6
番茄果实横茎(a)、纵茎(b)的模拟与实测散点图
Fig.6
Scatter graph of simulated and measured tomato fruit width (a) and length (b)
图7
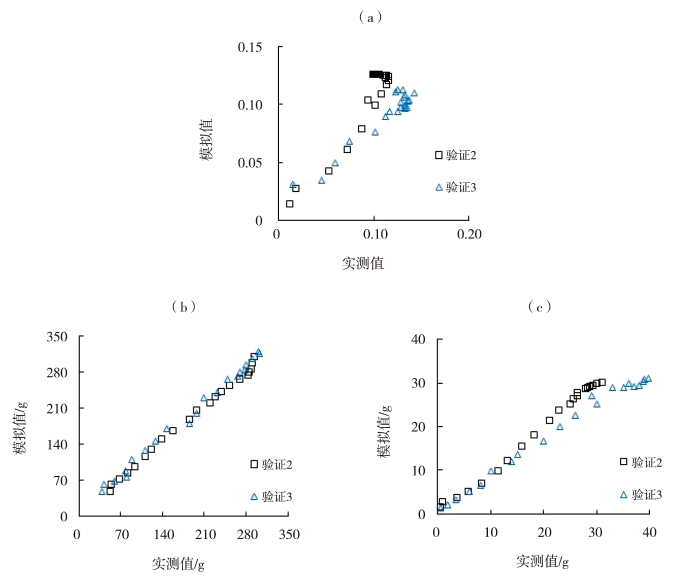
图7
番茄果实分配比(a)、总干物质量(b)、果实干物质量(c)的模拟与实测散点图
Fig.7
Scatter graph of simulated and measured fruit matter partitioning (a), total dry weight (b), and fruit dry weight (c) of tomato
表4 模型检验结果
Tab.4
| 验证 | R2 | RMSE | nRMSE | |
|---|---|---|---|---|
| 开花/朵 | 1 | 0.970 | 0.728 | 0.196 |
| 2 | 0.966 | 0.780 | 0.235 | |
| 3 | 0.839 | 0.753 | 0.246 | |
| 坐果/个 | 1 | 0.965 | 0.564 | 0.147 |
| 2 | 0.997 | 0.208 | 0.061 | |
| 3 | 0.974 | 0.588 | 0.145 | |
| 横茎/mm | 1 | 0.987 | 4.246 | 0.080 |
| 2 | 0.993 | 2.690 | 0.053 | |
| 3 | 0.992 | 7.436 | 0.133 | |
| 纵茎/mm | 1 | 0.999 | 2.932 | 0.078 |
| 2 | 0.989 | 0.991 | 0.027 | |
| 3 | 0.988 | 6.182 | 0.147 | |
| 果实分配比 | 2 | 0.905 | 0.015 | 0.164 |
| 3 | 0.925 | 0.026 | 0.227 | |
| 总干物质量/g | 2 | 0.996 | 6.435 | 0.035 |
| 3 | 0.993 | 14.229 | 0.078 | |
| 果实干物质量/g | 2 | 0.993 | 0.878 | 0.046 |
| 3 | 0.989 | 4.893 | 0.213 |
3 讨论与结论
3.1 讨论
辐热积模型综合考虑了温度和光照对作物的影响,通过选择作物的三基点温度中的最适生长范围温度,构建了光温与作物生长发育间的非线性关系。倪纪恒等(2009)发现,基于辐热积的Logistic曲线模型模拟温室黄瓜生长方面,精确度比使用有效积温模型提高了12.2%。因此,本文选用辐热积作为设施番茄生长的驱动因子建立Logistic模型曲线。随着辐热积的增加,番茄的生长进程呈现“S”型曲线。与以往研究(李永秀等,2006;倪纪恒,2009;石小虎等,2016;刘福昊等,2022)相比,本文增加了对番茄开花数、坐果数与果茎生长量等指标的模拟,使得模拟更为系统和细致。使用不同茬口、不同种植地区和品种的设施番茄对模型进行验证,进一步证实了模型的精确性与普适性。番茄开花数、坐果数和果实生长量等指标随辐热积增长到一定程度时,生长速率达到最大。因此,在番茄的最适温度范围内,提高温度或光照可增加辐热积积累,从而提高番茄的生长速率,促进其成熟。
研究结果表明,番茄生长指标模型的拟合度高、预测结果精确。在对开花数的验证中,不同茬口番茄生长指标模型的精确度最高,原因可能是同一番茄植株的生长状态和生育环境最为相似,从而导致不同茬口间差异最小。在验证1中,番茄的开花数和坐果数的实测值均高于预测值,可能因为第三茬花序开始生长时,番茄植株进入快速生长阶段,致使开花坐果速率较模型预测有所提高,而当番茄进入坐果期后,整体植株生长速率再次加快,致使第三茬番茄果实的生长速率与模型预测的差异增大。通过分析干物质分配的验证结果可以发现,在相似的生长环境下,不同品种间的差异远大于其他因素,尤其是在试验3中精确度最低。
本文的观测在设施作物生长适宜环境中进行,期间未遭遇长时间寡照灾害天气。尽管番茄植株在面对低温寡照环境时具有一定的自我调节能力,短期寡照不会破坏番茄的生长机理或影响最终产量(郁继华等,2004),但实际生长环境中若遭遇长时间低温寡照,超出了番茄自身调节能力的情况下,使用本模型可能会产生一定的误差。未来研究还需进一步完善,建立在不同程度低温寡照灾害后的辐热积生长模型。
3.2 结论
通过日光温室大棚中进行设施番茄生长试验,以辐热积为自变量、番茄生长指标为因变量构建Logistic曲线模型,然后利用不同茬口、品种、种植地区的番茄生长数据对模型进行验证,得到如下主要结论。
1)番茄开花数、坐果数和果实长成需要的累计辐热积分别为146.59、146.90、252.00 mol·m-2;当累计辐热积为62.19 mol·m-2时,番茄总干物质积累速率最大,为1.01 g·d-1;当累计辐热积为53.35 mol·m-2时,番茄花果干物质积累速率最大,为0.16 g·d-1。
2)番茄茎叶和根的分配比与辐热积的关系不适于用logistic曲线方程描述。番茄开花坐果数、果茎生长量、果实分配比与干物质重的模拟值与实测值的RMSE为0.208~14.229,nRMSE为0.027~0.246,R2为0.905~0.999,精确度整体较高,拟合效果较好。
3)不同地区和品种对番茄生长模型的精确度影响较大,而不同茬口的差异较小。模型验证显示,尽管不同地区、品种和茬口的精确度有所不同,模型的拟合度普遍较高,证明了模型的适用性。
参考文献
河南省设施农业冬季低温灾害风险评估
[J].
为了提高气候资源利用率,合理优化河南省冬季设施农业布局,利用1981—2018年河南省113个国家气象站冬季逐日最低气温资料,采用数理统计方法计算冬季年最低气温标准差和不同低温灾害指标的气候概率、发生频率、发生强度,并采用等权重法构建设施农业低温风险指数,结合低温灾情资料,选用有序样本最优聚类法划分设施农业低温风险等级,且叠加地表覆盖数据进行低温灾害风险评估。结果表明:河南省各地冬季低温灾害指标小于等于-5 ℃的气候概率和发生频率均超过0.80,不适宜发展塑料小拱棚,且不宜采用单层塑料大棚进行设施农业生产。信阳和南阳南部小于等于-10 ℃的气候概率和发生频率均小于0.20,适宜发展塑料大棚;鹤壁、安阳和濮阳小于等于-10 ℃的气候概率和发生频率均超过0.80,需发展日光温室才能保证设施作物生长。在塑料大棚发展区内,信阳市固始县和商城县为低温灾害轻度风险区,信阳大部、南阳部分区域和周口为中度风险区,其他地区均为重度风险区;在日光温室发展区内,濮阳、安阳和鹤壁部分区域为低温灾害特重风险区。河南省冬季各地不同等级的低温灾害风险有一定的区域性,在发展设施农业时,应因地制宜进行科学布局。
基于辐射和温度热效应的温室水果黄瓜叶面积模型
[J].依据温室黄瓜(Cucumis sativus)叶片生长与温度和辐射的关系,构建了适合我国种植技术的黄瓜叶面积模拟模型,并利用不同品种、播期的试验资料对模型进行了检验。结果表明,该模型比传统的积温法和比叶面积法更准确地模拟温室水果黄瓜的叶面积。该模型对黄瓜叶面积指数的模拟结果与1∶1直线之间的决定系数R2和回归估计标准误差RMSE分别为0.879 2和0.398 0,比用积温法和比叶面积法模拟叶面积指数的精度分别提高了37%和74%。
A simple phenological model of muskmelon development
[J].
An extension of the logistic model of plant growth
[J].
Modelling phasic development in wheat: a conceptual integration of physiological components
[J].
Quantifying the responses to temperature of different plant processes involved in growth and development of glasshouse tomato
[J].
Using sigmoid growth models to simulate greenhouse tomato growth and development
[J].
Modeling the relationship between tomato fruit growth and the effective accumulated temperature in solar greenhouse
[J].
Growth, development and yield of a tomato crop: periodic destructive measurements in a greenhouse
[J].
Evaluation of a dynamic simulation model for tomato crop growth and development
[J].
A dynamic tomato growth and yield model (tomgro)
[J].
Temperature driven leaf unfolding rate in Hibiscus rosa-sinensis
[J].
Simulation of biomass allocation in greenhouse crops-a review
[J].
Fruit growth and biomass allocation to the fruits in cucumber. 1. Effect of fruit load and temperature
[J].
A simulation model for dry matter partitioning in cucumber
[J].A dynamic model is developed for the simulation of the daily dry matter distribution between the generative and vegetative plant parts and the distribution among individual fruits in greenhouse cucumber. The model is based on the hypothesis that dry matter partitioning is regulated by the sink strengths of the plant organs. The sink strength of an organ is denned here as its potential growth rate, i.e. the growth rate at non-limiting assimilate supply. The sink strength of each individual fruit is described as a function of its temperature sum after anthesis and the actual temperature, that of the vegetative plant parts as a function of actual temperature only. The formation rate of nonaborting fruits is essentially a function of the source/sink ratio. Model results agreed well with the measured fluctuating distribution of dry matter between fruits and vegetative parts. The measured effects of three intensities of fruit removal were also simulated satisfactorily. When simulating the partitioning among individual fruits the final fruit size was simulated quite well. However, the growth rate of young fruits was usually overestimated and that of old fruits underestimated, because of dominance among fruits. This phenomenon could be accounted for by incorporating priority functions into the model. Finally, a sensitivity analysis of the model was performed to investigate the effects of some climatic factors, manipulations of the number of fruits on a plant and model parameters on dry matter distribution. Strategies to manipulate the dry matter distribution are discussed.
Photosystem inhibition and protection in tomato leaves under low light
[J].
On temperature and water limitation of net ecosystem productivity: Implementation in the C-Fix model
[J].
Crop continuity and prediction of maturity in the crisp lettuce variety Saladin
[J].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}